

The commonness of rarity: Global and future distribution of rarity across land plants
- PMID: 31807712
- PMCID: PMC6881168
- DOI: 10.1126/sciadv.aaz0414
The commonness of rarity: Global and future distribution of rarity across land plants
Abstract
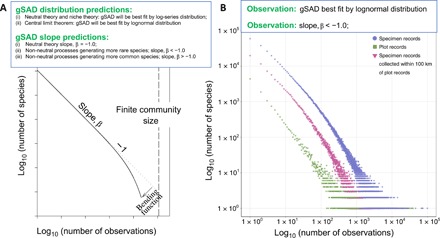
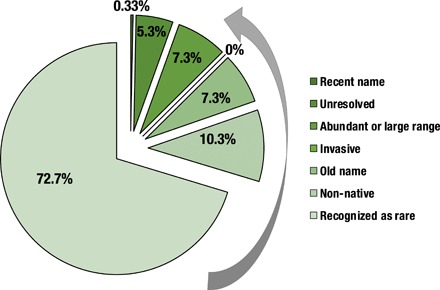
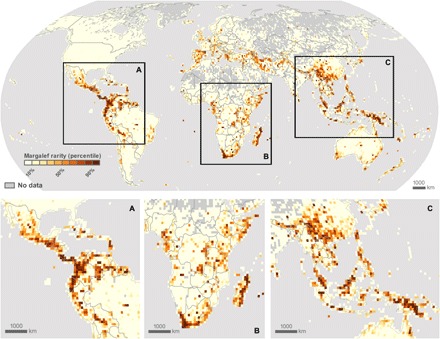
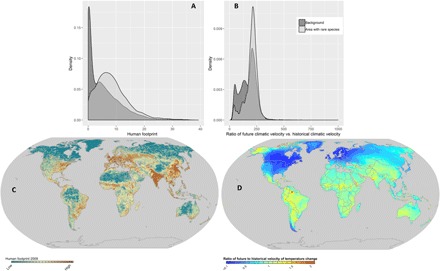
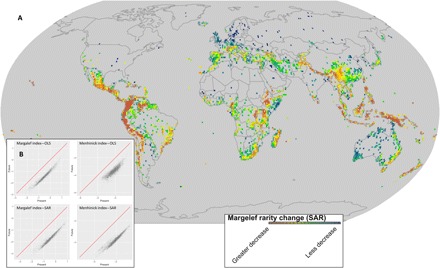
A key feature of life's diversity is that some species are common but many more are rare. Nonetheless, at global scales, we do not know what fraction of biodiversity consists of rare species. Here, we present the largest compilation of global plant diversity to quantify the fraction of Earth's plant biodiversity that are rare. A large fraction, ~36.5% of Earth's ~435,000 plant species, are exceedingly rare. Sampling biases and prominent models, such as neutral theory and the k-niche model, cannot account for the observed prevalence of rarity. Our results indicate that (i) climatically more stable regions have harbored rare species and hence a large fraction of Earth's plant species via reduced extinction risk but that (ii) climate change and human land use are now disproportionately impacting rare species. Estimates of global species abundance distributions have important implications for risk assessments and conservation planning in this era of rapid global change.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
Similar articles
-
Rarity in freshwater vascular plants across Europe and North America: Patterns, mechanisms and future scenarios.Sci Total Environ. 2021 Sep 10;786:147491. doi: 10.1016/j.scitotenv.2021.147491. Epub 2021 May 3. Sci Total Environ. 2021. PMID: 33965814
-
Fire and biodiversity in the Anthropocene.Science. 2020 Nov 20;370(6519):eabb0355. doi: 10.1126/science.abb0355. Science. 2020. PMID: 33214246 Review.
-
Future habitat loss and extinctions driven by land-use change in biodiversity hotspots under four scenarios of climate-change mitigation.Conserv Biol. 2015 Aug;29(4):1122-1131. doi: 10.1111/cobi.12549. Epub 2015 Jun 30. Conserv Biol. 2015. PMID: 26129841
-
Global change biology: A primer.Glob Chang Biol. 2020 Jan;26(1):3-30. doi: 10.1111/gcb.14893. Epub 2019 Dec 9. Glob Chang Biol. 2020. PMID: 31663217 Review.
-
Predicting loss of evolutionary history: Where are we?Biol Rev Camb Philos Soc. 2017 Feb;92(1):271-291. doi: 10.1111/brv.12228. Epub 2015 Oct 14. Biol Rev Camb Philos Soc. 2017. PMID: 26467982 Review.
Cited by
-
Widespread homogenization of plant communities in the Anthropocene.Nat Commun. 2021 Dec 6;12(1):6983. doi: 10.1038/s41467-021-27186-8. Nat Commun. 2021. PMID: 34873159 Free PMC article.
-
Global patterns and ecological drivers of taxonomic and phylogenetic endemism in angiosperm genera.Plant Divers. 2024 Jan 4;46(2):149-157. doi: 10.1016/j.pld.2023.11.004. eCollection 2024 Mar. Plant Divers. 2024. PMID: 38807907 Free PMC article.
-
Consistent patterns of common species across tropical tree communities.Nature. 2024 Jan;625(7996):728-734. doi: 10.1038/s41586-023-06820-z. Epub 2024 Jan 10. Nature. 2024. PMID: 38200314 Free PMC article.
-
Navigating transformation of biodiversity and climate.Sci Adv. 2019 Nov 27;5(11):eaba0969. doi: 10.1126/sciadv.aba0969. eCollection 2019 Nov. Sci Adv. 2019. PMID: 31832538 Free PMC article. No abstract available.
-
Climate change and land use threaten global hotspots of phylogenetic endemism for trees.Nat Commun. 2023 Oct 31;14(1):6950. doi: 10.1038/s41467-023-42671-y. Nat Commun. 2023. PMID: 37907453 Free PMC article.
References
-
- Preston F. W., The canonical distribution of commonness and rarity: Part I. Ecology 43, 185–215 (1962).
-
- Hanski I., Dynamics of regional distribution: The core and satellite species hypothesis. Oikos 38, 210–221 (1982).
-
- J. M. Diamond, “Normal” extinctions of isolated populations, in Extinctions, M. H. Nitecki, Ed. (University of Chicago Press, 1984), pp. 191–246.
-
- Pimm S. L., Jones H. L., Diamond J., On the risk of extinction. Am. Nat. 132, 757–785 (1988).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Research Materials
