

Hic1 Defines Quiescent Mesenchymal Progenitor Subpopulations with Distinct Functions and Fates in Skeletal Muscle Regeneration
- PMID: 31809738
- PMCID: PMC6941576
- DOI: 10.1016/j.stem.2019.11.004
Hic1 Defines Quiescent Mesenchymal Progenitor Subpopulations with Distinct Functions and Fates in Skeletal Muscle Regeneration
Abstract
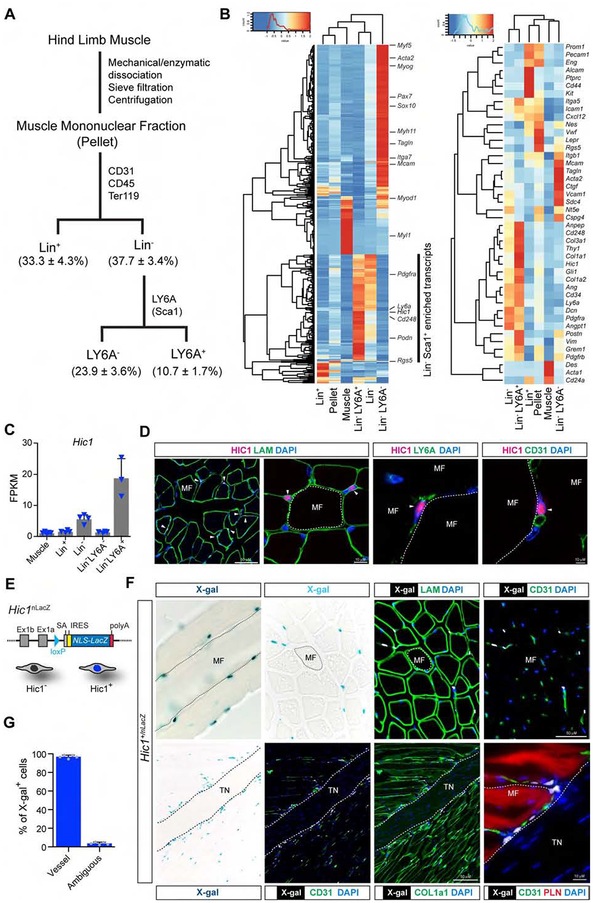
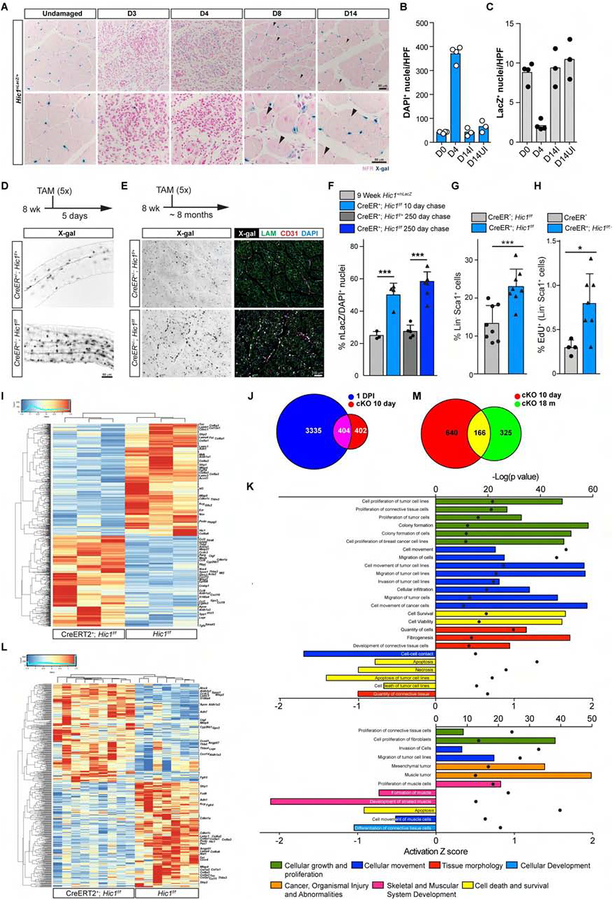
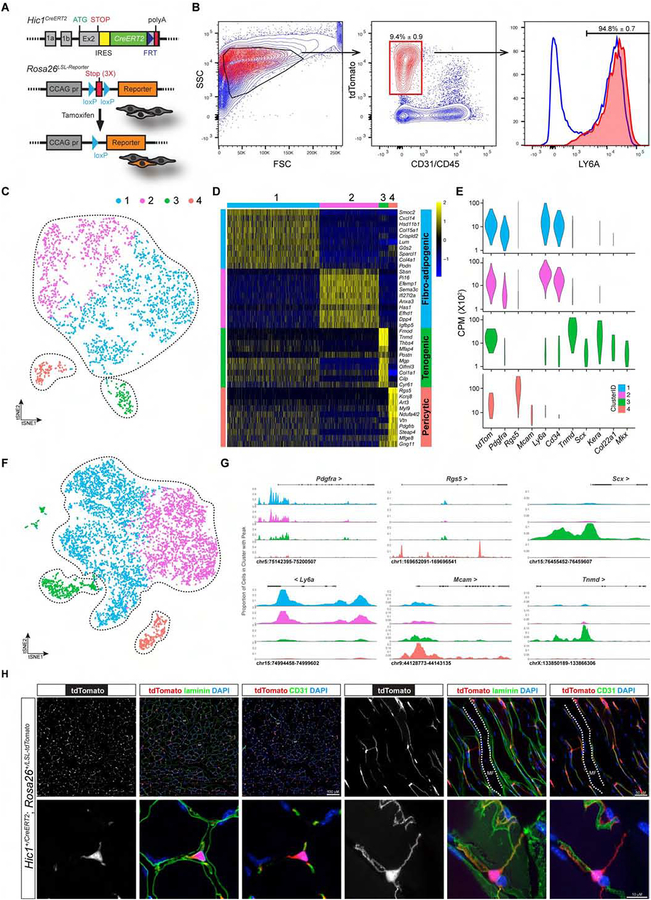
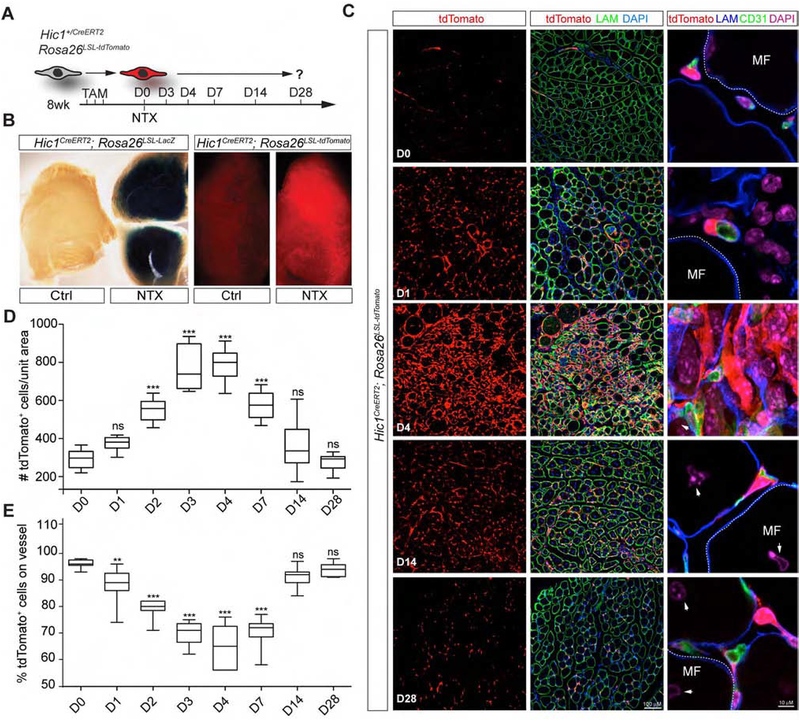
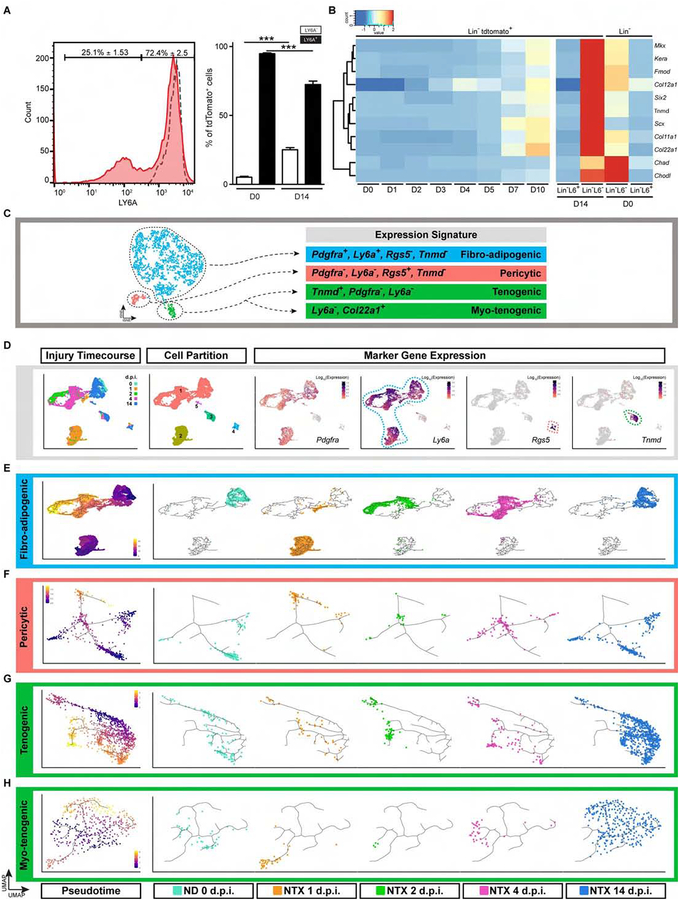
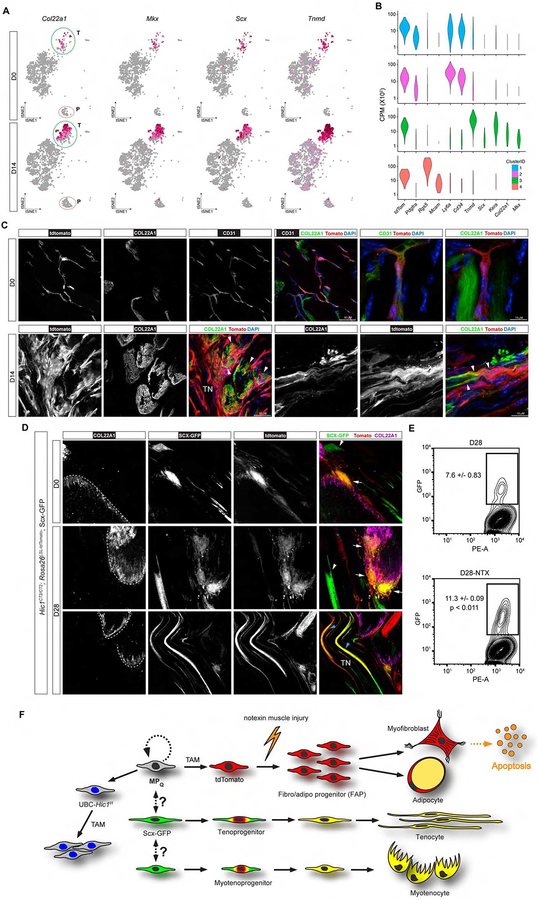
Many adult tissues contain resident stem cells, such as the Pax7+ satellite cells within skeletal muscle, that regenerate parenchymal elements following damage. Tissue-resident mesenchymal progenitors (MPs) also participate in regeneration, although their function and fate in this process are unclear. Here, we identify Hypermethylated in cancer 1 (Hic1) as a marker of MPs in skeletal muscle and further show that Hic1 deletion leads to MP hyperplasia. Single-cell RNA-seq and ATAC-seq analysis of Hic1+ MPs in skeletal muscle shows multiple subpopulations, which we further show have distinct functions and lineage potential. Hic1+ MPs orchestrate multiple aspects of skeletal muscle regeneration by providing stage-specific immunomodulation and trophic and mechanical support. During muscle regeneration, Hic1+ derivatives directly contribute to several mesenchymal compartments including Col22a1-expressing cells within the myotendinous junction. Collectively, these findings demonstrate that HIC1 regulates MP quiescence and identifies MP subpopulations with transient and enduring roles in muscle regeneration.
Keywords: lineage tracing; mesenchymal progenitors; myotendinous junction; pericytes; quiescence; scATAC-seq; scRNA-seq; skeletal muscle; tendon; tissue regeneration.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors have no competing interests to declare.
Figures

Comment in
-
Keeping Fibrotic Responses in Contractile Tissues at Bay: The Plot t(Hic1)ens.Cell Stem Cell. 2020 Feb 6;26(2):129-130. doi: 10.1016/j.stem.2019.12.010. Cell Stem Cell. 2020. PMID: 32032520
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
