

Irregular Chromatin: Packing Density, Fiber Width, and Occurrence of Heterogeneous Clusters
- PMID: 31810656
- PMCID: PMC6950648
- DOI: 10.1016/j.bpj.2019.11.004
Irregular Chromatin: Packing Density, Fiber Width, and Occurrence of Heterogeneous Clusters
Abstract
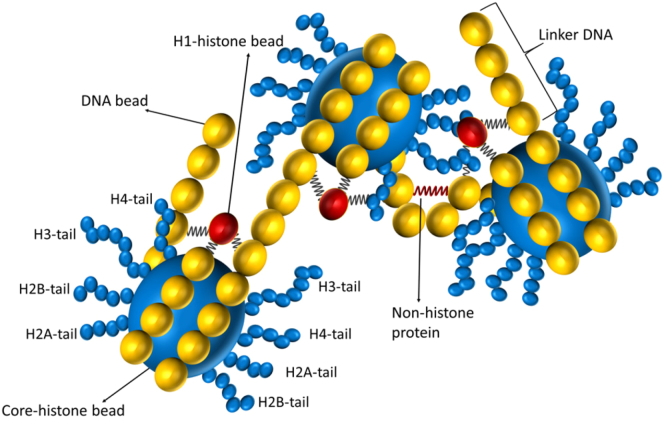
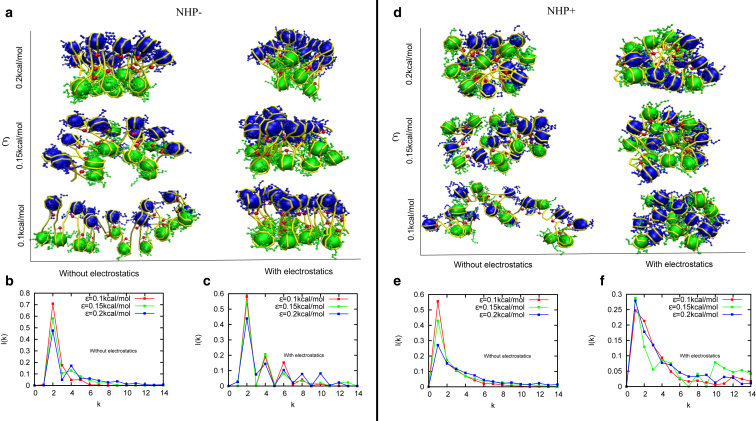
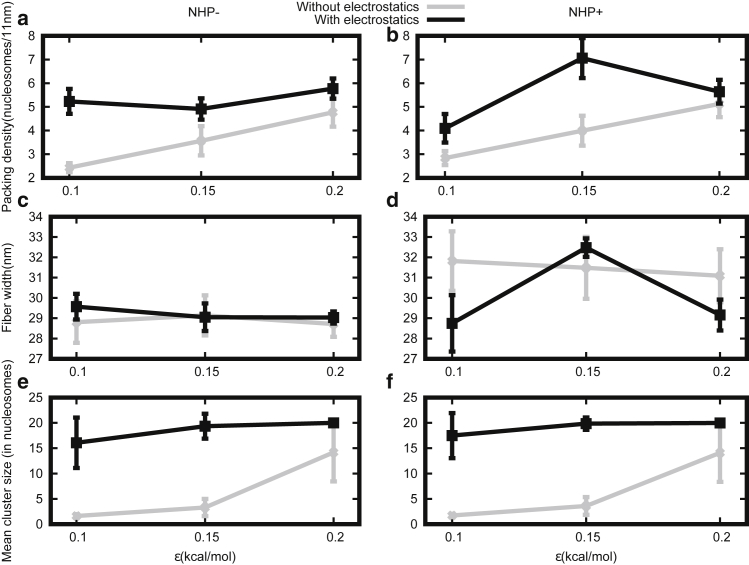
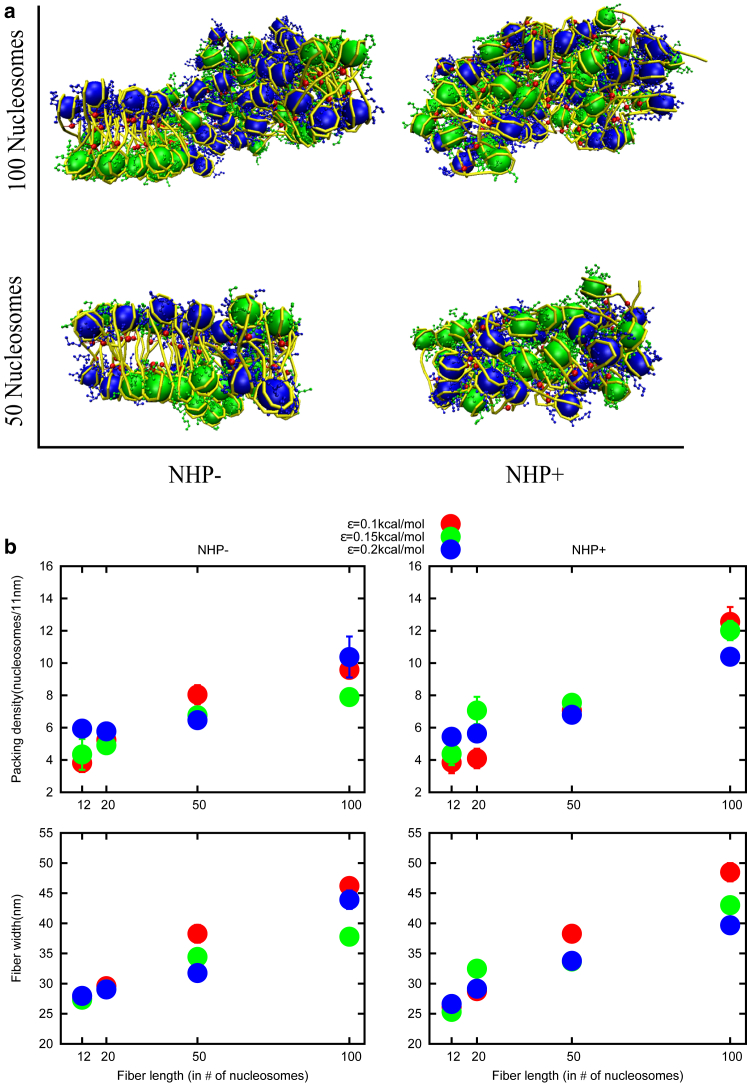
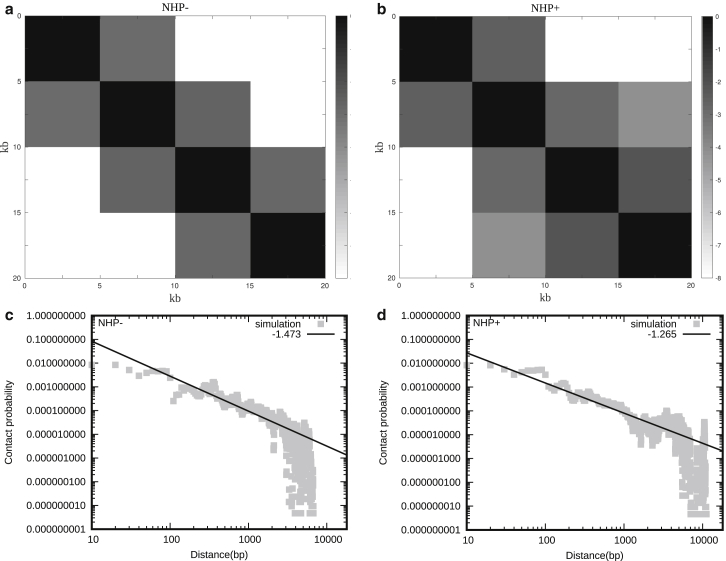
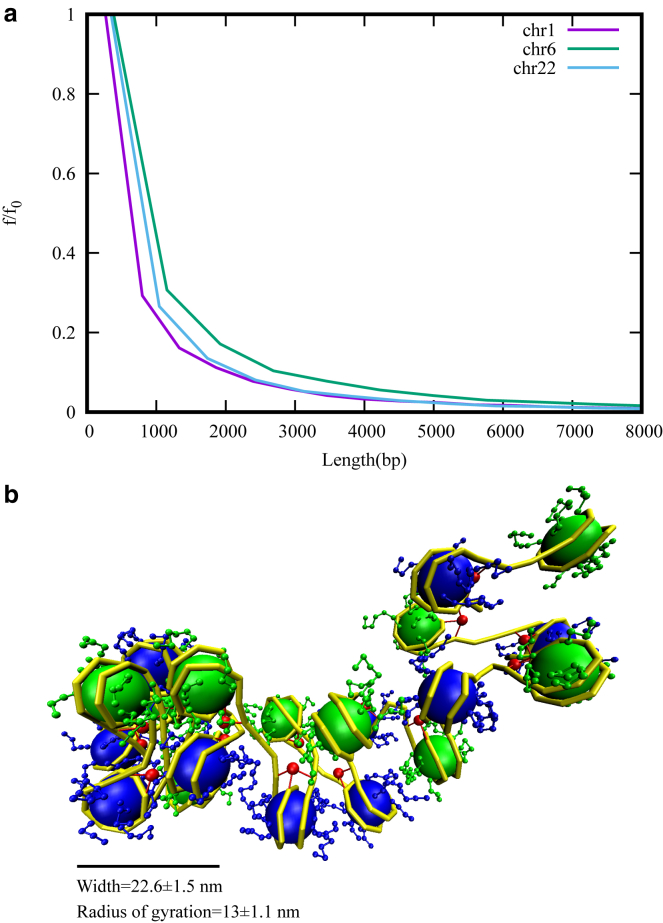
How chromatin is folded on the length scale of a gene is an open question. Recent experiments have suggested that, in vivo, chromatin is folded in an irregular manner and not as an ordered fiber with a width of 30 nm that is expected from theories of higher order packaging. Using computational methods, we examine how the interplay between DNA-bending nonhistone proteins, histone tails, intrachromatin electrostatic, and other interactions decide the nature of the packaging of chromatin. We show that although the DNA-bending nonhistone proteins make the chromatin irregular, they may not alter the packing density and size of the fiber. We find that the length of the interacting region and intrachromatin electrostatic interactions influence the packing density, clustering of nucleosomes, and the width of the chromatin fiber. Our results suggest that the heterogeneity in the interaction pattern will play an important role in deciding the nature of the packaging of chromatin.
Copyright © 2019 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Alberts B. Sixth Edition. Garland Science, Taylor and Francis Group; New York: 2014. Molecular Biology of the Cell.
-
- Kornberg R.D. Chromatin structure: a repeating unit of histones and DNA. Science. 1974;184:868–871. - PubMed
-
- Luger K., Mäder A.W., Richmond T.J. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature. 1997;389:251–260. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
