

The local translation of KNa in dendritic projections of auditory neurons and the roles of KNa in the transition from hidden to overt hearing loss
- PMID: 31812952
- PMCID: PMC6932877
- DOI: 10.18632/aging.102553
The local translation of KNa in dendritic projections of auditory neurons and the roles of KNa in the transition from hidden to overt hearing loss
Abstract
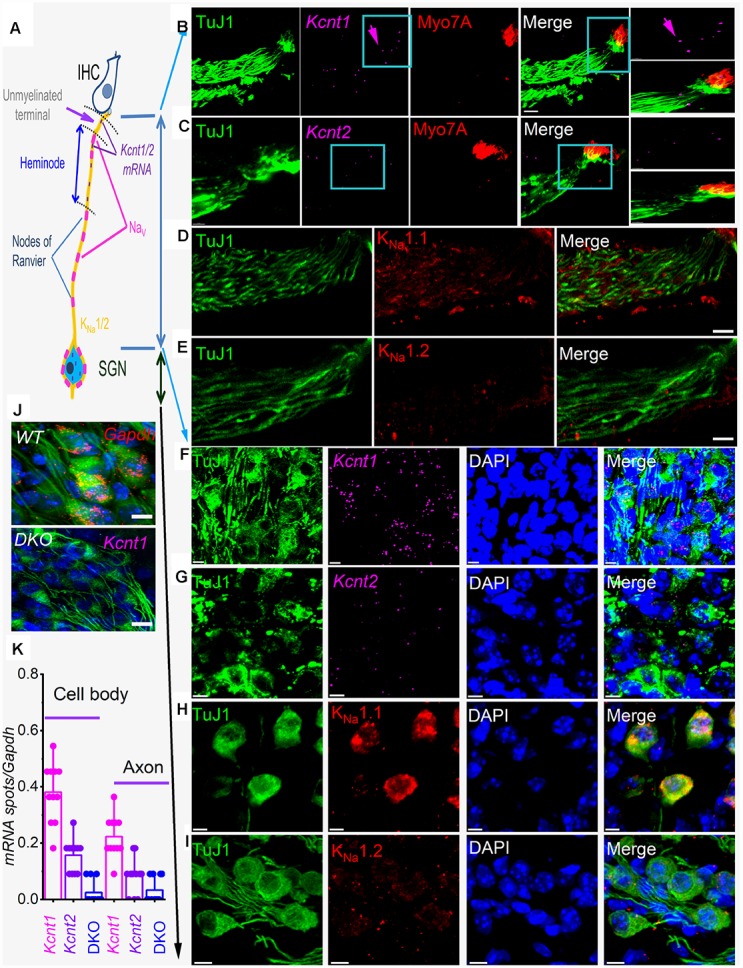
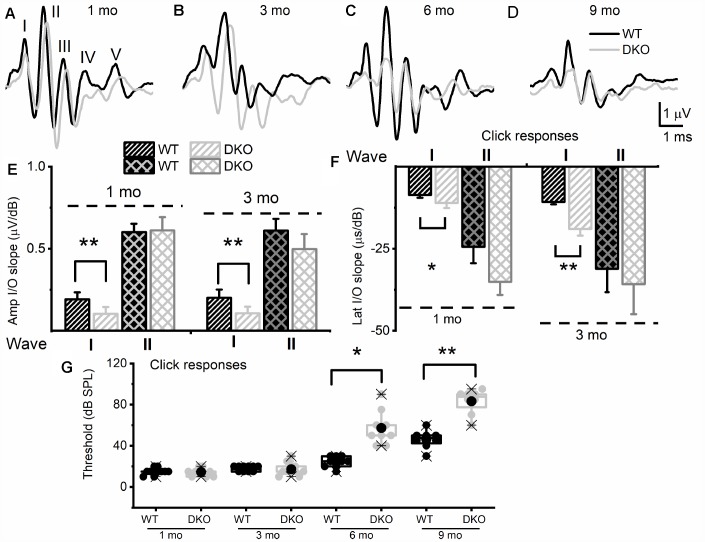
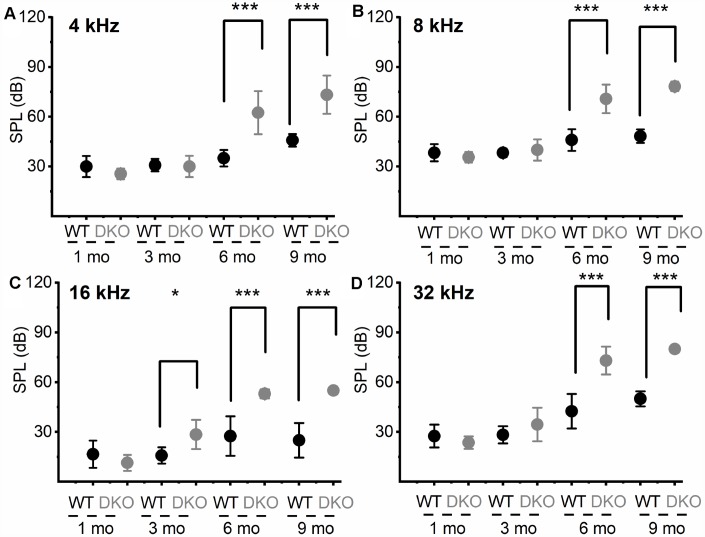
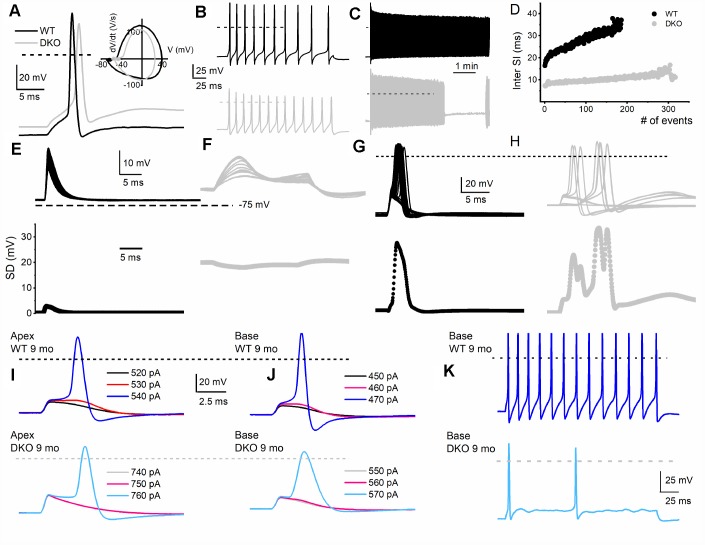
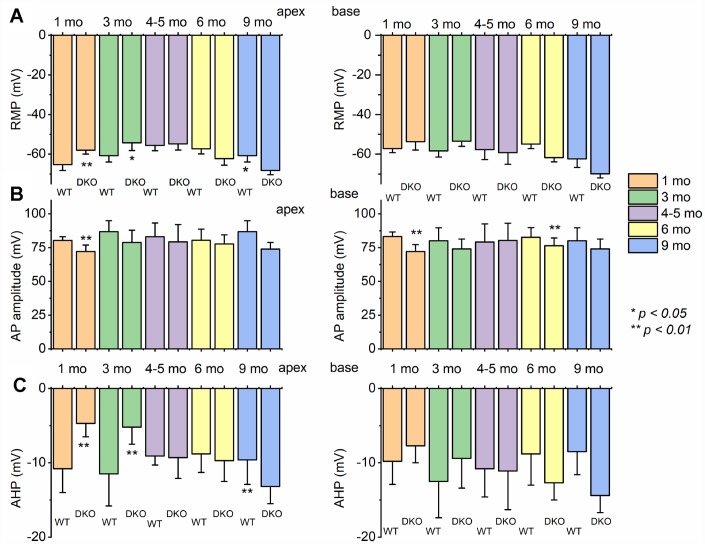
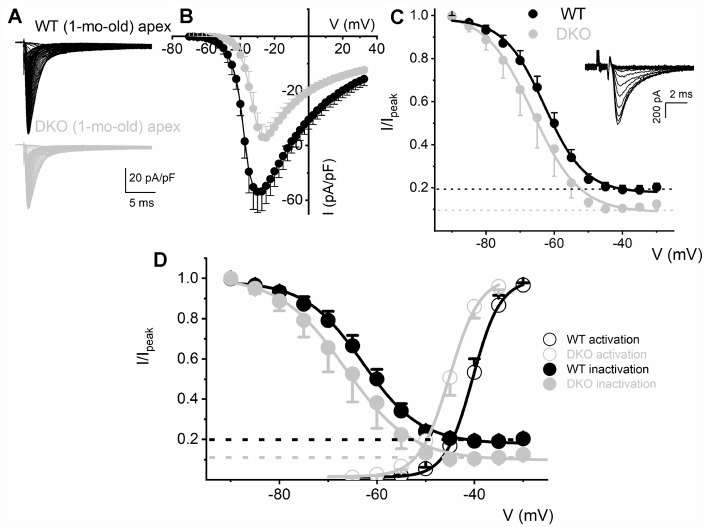
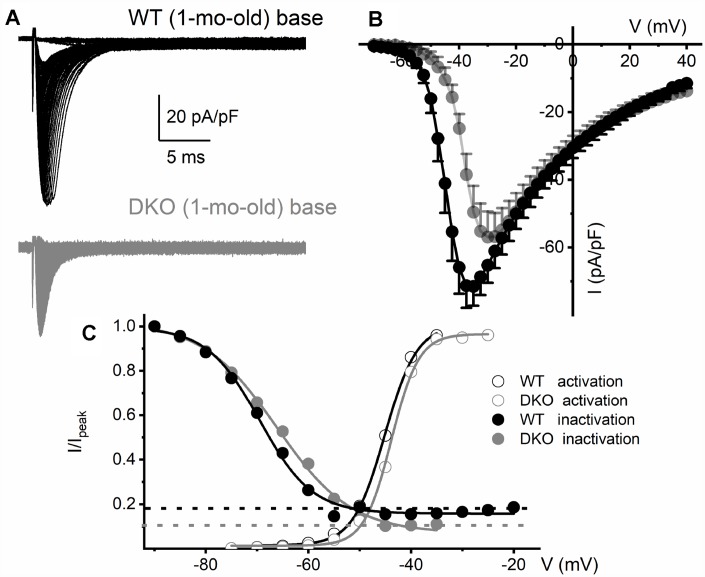
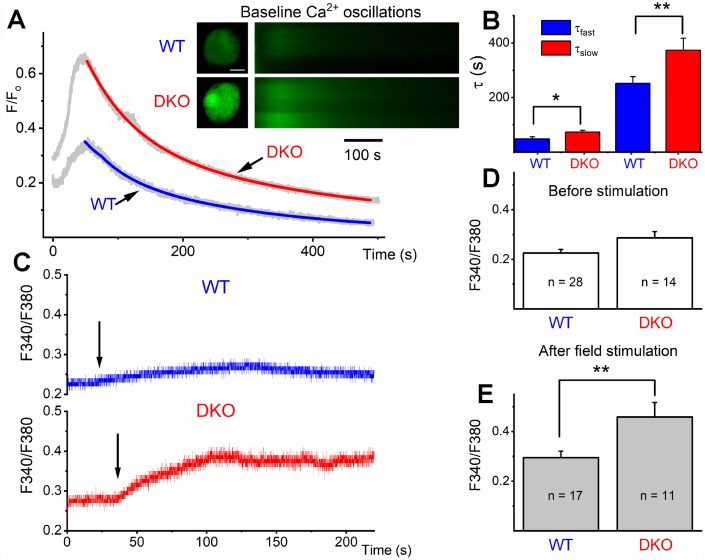
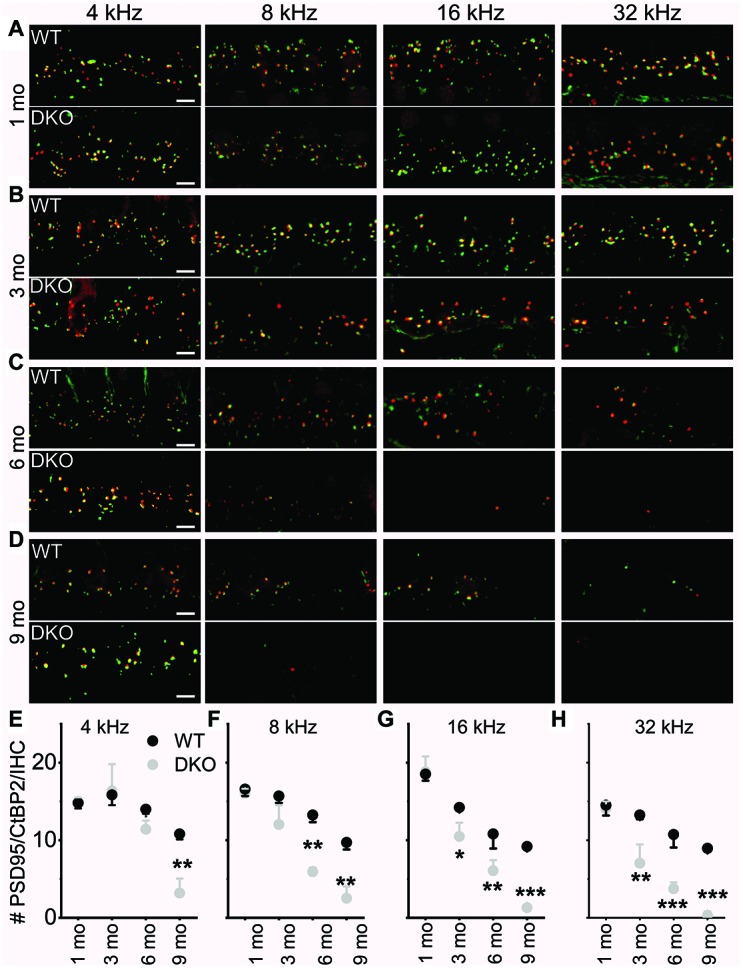
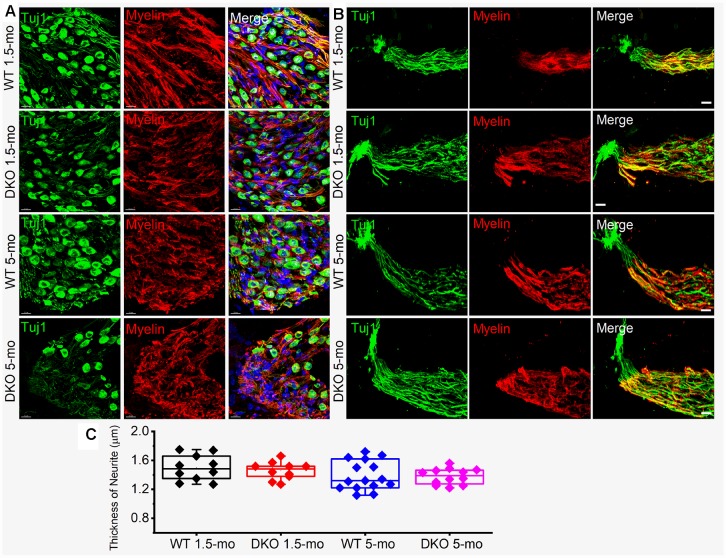
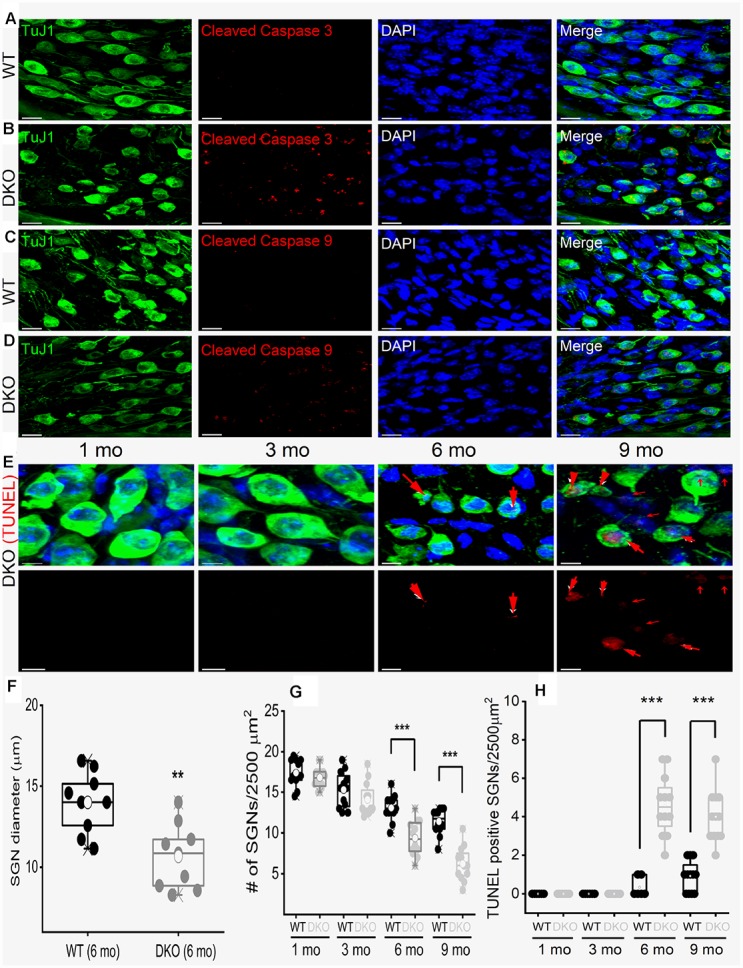
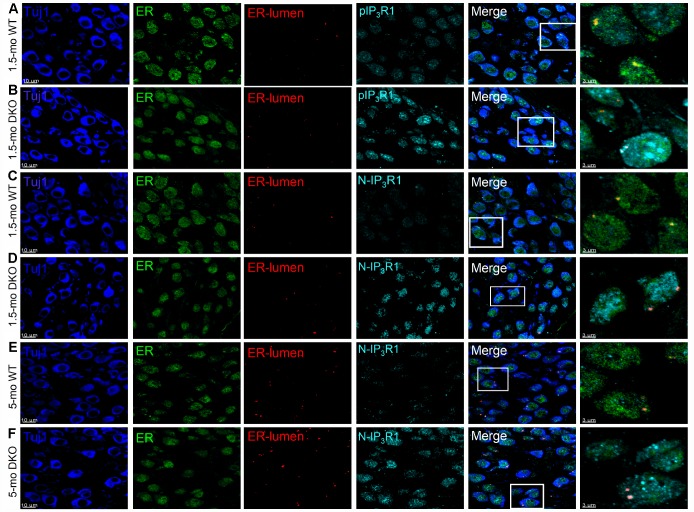
Local and privileged expression of dendritic proteins allows segregation of distinct functions in a single neuron but may represent one of the underlying mechanisms for early and insidious presentation of sensory neuropathy. Tangible characteristics of early hearing loss (HL) are defined in correlation with nascent hidden hearing loss (HHL) in humans and animal models. Despite the plethora of causes of HL, only two prevailing mechanisms for HHL have been identified, and in both cases, common structural deficits are implicated in inner hair cell synapses, and demyelination of the auditory nerve (AN). We uncovered that Na+-activated K+ (KNa) mRNA and channel proteins are distinctly and locally expressed in dendritic projections of primary ANs and genetic deletion of KNa channels (Kcnt1 and Kcnt2) results in the loss of proper AN synaptic function, characterized as HHL, without structural synaptic alterations. We further demonstrate that the local functional synaptic alterations transition from HHL to increased hearing-threshold, which entails changes in global Ca2+ homeostasis, activation of caspases 3/9, impaired regulation of inositol triphosphate receptor 1 (IP3R1), and apoptosis-mediated neurodegeneration. Thus, the present study demonstrates how local synaptic dysfunction results in an apparent latent pathological phenotype (HHL) and, if undetected, can lead to overt HL. It also highlights, for the first time, that HHL can precede structural synaptic dysfunction and AN demyelination. The stepwise cellular mechanisms from HHL to canonical HL are revealed, providing a platform for intervention to prevent lasting and irreversible age-related hearing loss (ARHL).
Keywords: age-related hearing loss; auditory neurons; axonal protein translation; hearing loss; potassium channels.
Conflict of interest statement
Figures
Similar articles
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
Sodium-activated potassium channels shape peripheral auditory function and activity of the primary auditory neurons in mice.Sci Rep. 2019 Feb 22;9(1):2573. doi: 10.1038/s41598-019-39119-z. Sci Rep. 2019. PMID: 30796290 Free PMC article.
-
Prescription of Controlled Substances: Benefits and Risks.2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jul 6. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 30726003 Free Books & Documents.
-
COL1A1- and COL1A2-Related Osteogenesis Imperfecta.2005 Jan 28 [updated 2025 May 29]. In: Adam MP, Feldman J, Mirzaa GM, Pagon RA, Wallace SE, Amemiya A, editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993–2025. 2005 Jan 28 [updated 2025 May 29]. In: Adam MP, Feldman J, Mirzaa GM, Pagon RA, Wallace SE, Amemiya A, editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993–2025. PMID: 20301472 Free Books & Documents. Review.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
Cited by
-
Guanylyl Cyclase A/cGMP Signaling Slows Hidden, Age- and Acoustic Trauma-Induced Hearing Loss.Front Aging Neurosci. 2020 Apr 9;12:83. doi: 10.3389/fnagi.2020.00083. eCollection 2020. Front Aging Neurosci. 2020. PMID: 32327991 Free PMC article.
-
Exploring the potential biomarkers and potential causality of Ménière disease based on bioinformatics and machine learning.Medicine (Baltimore). 2025 May 9;104(19):e42399. doi: 10.1097/MD.0000000000042399. Medicine (Baltimore). 2025. PMID: 40355226 Free PMC article.
-
KCNT1 gene variant-associated epilepsy: genetic insights, functional mechanisms, and emerging therapies.J Neurol. 2025 Jun 21;272(7):472. doi: 10.1007/s00415-025-13207-9. J Neurol. 2025. PMID: 40544226 Review.
-
Prestin amplifies cardiac motor functions.Cell Rep. 2021 May 4;35(5):109097. doi: 10.1016/j.celrep.2021.109097. Cell Rep. 2021. PMID: 33951436 Free PMC article.
-
Sodium sensitivity of KNa channels in mouse CA1 neurons.J Neurophysiol. 2021 May 1;125(5):1690-1697. doi: 10.1152/jn.00064.2021. Epub 2021 Mar 31. J Neurophysiol. 2021. PMID: 33788620 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
