

Ketamine Restores Thalamic-Prefrontal Cortex Functional Connectivity in a Mouse Model of Neurodevelopmental Disorder-Associated 2p16.3 Deletion
- PMID: 31812984
- PMCID: PMC7175007
- DOI: 10.1093/cercor/bhz244
Ketamine Restores Thalamic-Prefrontal Cortex Functional Connectivity in a Mouse Model of Neurodevelopmental Disorder-Associated 2p16.3 Deletion
Abstract
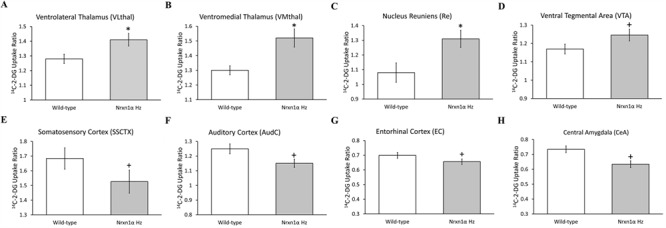
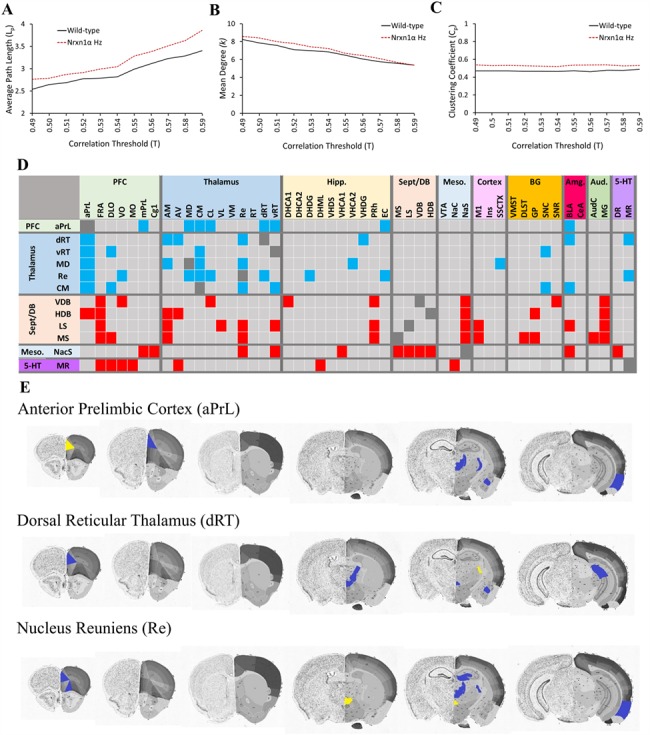
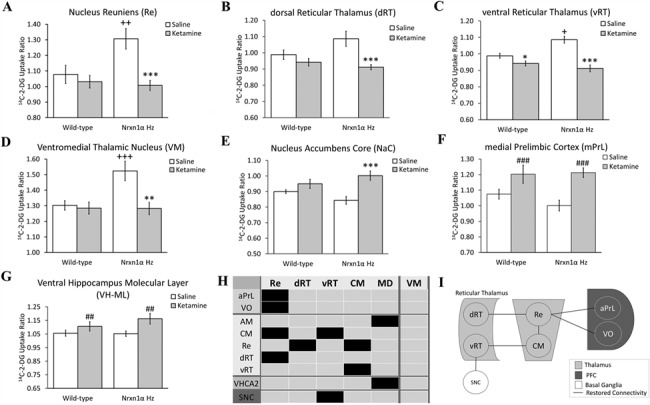
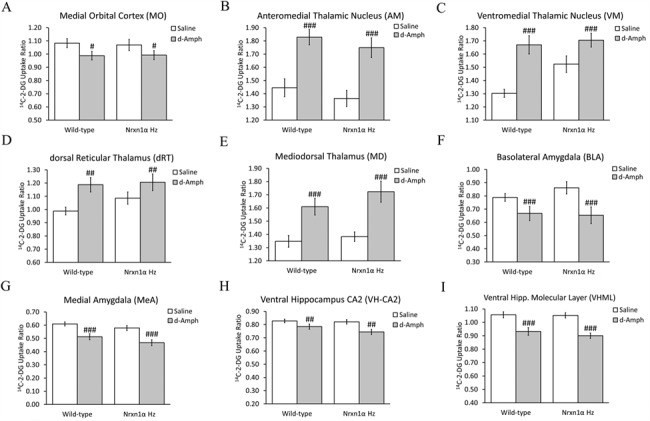
2p16.3 deletions, involving heterozygous NEUREXIN1 (NRXN1) deletion, dramatically increase the risk of developing neurodevelopmental disorders, including autism and schizophrenia. We have little understanding of how NRXN1 heterozygosity increases the risk of developing these disorders, particularly in terms of the impact on brain and neurotransmitter system function and brain network connectivity. Thus, here we characterize cerebral metabolism and functional brain network connectivity in Nrxn1α heterozygous mice (Nrxn1α+/- mice), and assess the impact of ketamine and dextro-amphetamine on cerebral metabolism in these animals. We show that heterozygous Nrxn1α deletion alters cerebral metabolism in neural systems implicated in autism and schizophrenia including the thalamus, mesolimbic system, and select cortical regions. Nrxn1α heterozygosity also reduces the efficiency of functional brain networks, through lost thalamic "rich club" and prefrontal cortex (PFC) hub connectivity and through reduced thalamic-PFC and thalamic "rich club" regional interconnectivity. Subanesthetic ketamine administration normalizes the thalamic hypermetabolism and partially normalizes thalamic disconnectivity present in Nrxn1α+/- mice, while cerebral metabolic responses to dextro-amphetamine are unaltered. The data provide new insight into the systems-level impact of heterozygous Nrxn1α deletion and how this increases the risk of developing neurodevelopmental disorders. The data also suggest that the thalamic dysfunction induced by heterozygous Nrxn1α deletion may be NMDA receptor-dependent.
Keywords: NMDA receptor; autism; functional brain imaging; graph theory; schizophrenia.
© The Author(s) 2019. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
Similar articles
-
Altered medial prefrontal cortex and dorsal raphé activity predict genotype and correlate with abnormal learning behavior in a mouse model of autism-associated 2p16.3 deletion.Autism Res. 2022 Apr;15(4):614-627. doi: 10.1002/aur.2685. Epub 2022 Feb 10. Autism Res. 2022. PMID: 35142069 Free PMC article.
-
Subanaesthetic ketamine treatment alters prefrontal cortex connectivity with thalamus and ascending subcortical systems.Schizophr Bull. 2013 Mar;39(2):366-77. doi: 10.1093/schbul/sbr144. Epub 2011 Nov 22. Schizophr Bull. 2013. PMID: 22114100 Free PMC article.
-
Bi-allelic NRXN1α deletion in microglia derived from iPSC of an autistic patient increases interleukin-6 production and impairs supporting function on neuronal networking.Brain Behav Immun. 2025 Jan;123:28-42. doi: 10.1016/j.bbi.2024.09.001. Epub 2024 Sep 5. Brain Behav Immun. 2025. PMID: 39243986
-
Phenotypic spectrum of NRXN1 mono- and bi-allelic deficiency: A systematic review.Clin Genet. 2020 Jan;97(1):125-137. doi: 10.1111/cge.13537. Epub 2019 May 7. Clin Genet. 2020. PMID: 30873608
-
Neural circuit dysfunction in mouse models of neurodevelopmental disorders.Curr Opin Neurobiol. 2018 Feb;48:174-182. doi: 10.1016/j.conb.2017.12.013. Epub 2018 Jan 10. Curr Opin Neurobiol. 2018. PMID: 29329089 Review.
Cited by
-
Ketamine Self-Medication in a Patient with Autism Spectrum Disorder and Comorbid Therapy-Resistant Depression.Psychiatry Clin Psychopharmacol. 2022 Sep 1;32(3):268-272. doi: 10.5152/pcp.2022.22037. eCollection 2022 Sep. Psychiatry Clin Psychopharmacol. 2022. PMID: 38766674 Free PMC article.
-
Ketamine impairs growth cone and synaptogenesis in human GABAergic projection neurons via GSK-3β and HDAC6 signaling.Mol Psychiatry. 2024 Jun;29(6):1647-1659. doi: 10.1038/s41380-022-01864-5. Epub 2022 Nov 21. Mol Psychiatry. 2024. PMID: 36414713 Free PMC article.
-
Allelic contribution of Nrxn1α to autism-relevant behavioral phenotypes in mice.PLoS Genet. 2023 Feb 27;19(2):e1010659. doi: 10.1371/journal.pgen.1010659. eCollection 2023 Feb. PLoS Genet. 2023. PMID: 36848371 Free PMC article.
-
Using Zebrafish to Model Autism Spectrum Disorder: A Comparison of ASD Risk Genes Between Zebrafish and Their Mammalian Counterparts.Front Mol Neurosci. 2020 Nov 11;13:575575. doi: 10.3389/fnmol.2020.575575. eCollection 2020. Front Mol Neurosci. 2020. PMID: 33262688 Free PMC article. Review.
-
Altered medial prefrontal cortex and dorsal raphé activity predict genotype and correlate with abnormal learning behavior in a mouse model of autism-associated 2p16.3 deletion.Autism Res. 2022 Apr;15(4):614-627. doi: 10.1002/aur.2685. Epub 2022 Feb 10. Autism Res. 2022. PMID: 35142069 Free PMC article.
References
-
- Alberi L, Lintas A, Kretz R, Schwaller B, Villa AEP. 2012. The calcium binding protein parvalbumin modulates the firing 1 properties of the reticular thalamus bursting neurons. J Neurophysiol. 109:2827–2841. - PubMed
-
- Aoki Y, Cortese S, Tansella M. 2015. Neural bases of atypical emotional face processing in autism: a meta-analysis of fMRI studies. World J Biol Psychiatry. 16:291–300. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
