

The majority of A-to-I RNA editing is not required for mammalian homeostasis
- PMID: 31815657
- PMCID: PMC6900863
- DOI: 10.1186/s13059-019-1873-2
The majority of A-to-I RNA editing is not required for mammalian homeostasis
Abstract
Background: Adenosine-to-inosine (A-to-I) RNA editing, mediated by ADAR1 and ADAR2, occurs at tens of thousands to millions of sites across mammalian transcriptomes. A-to-I editing can change the protein coding potential of a transcript and alter RNA splicing, miRNA biology, RNA secondary structure and formation of other RNA species. In vivo, the editing-dependent protein recoding of GRIA2 is the essential function of ADAR2, while ADAR1 editing prevents innate immune sensing of endogenous RNAs by MDA5 in both human and mouse. However, a significant proportion of A-to-I editing sites can be edited by both ADAR1 and ADAR2, particularly within the brain where both are highly expressed. The physiological function(s) of these shared sites, including those evolutionarily conserved, is largely unknown.
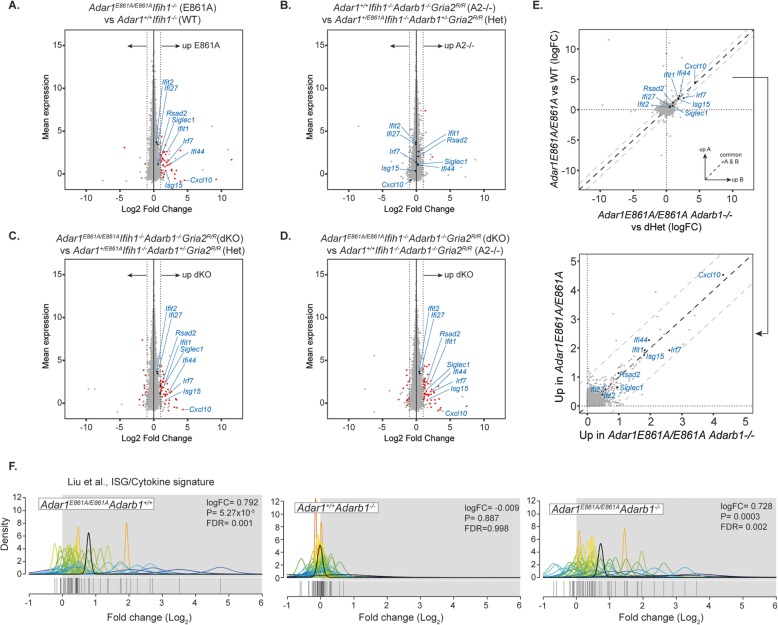
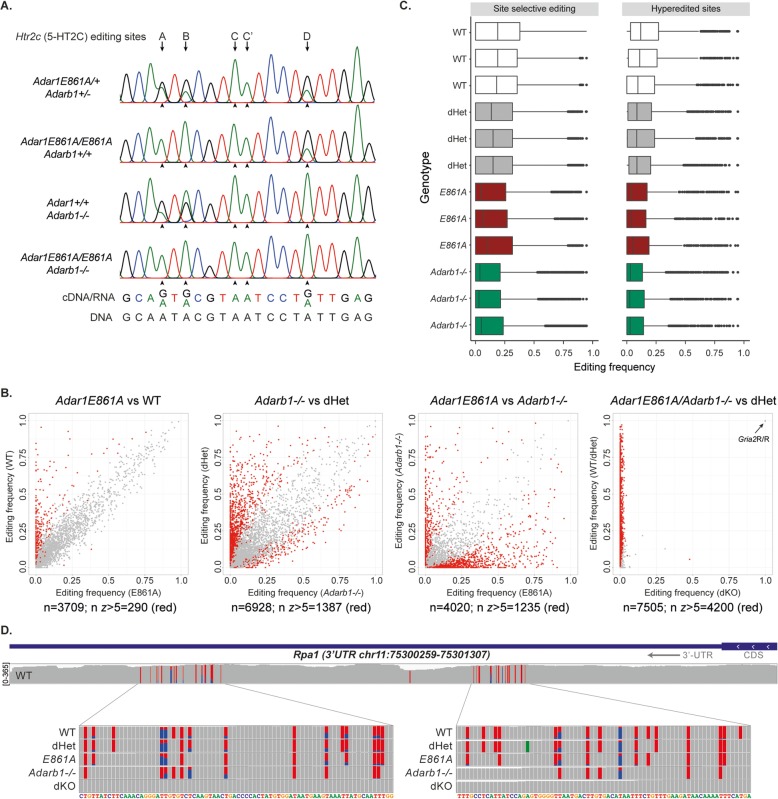
Results: To generate completely A-to-I editing-deficient mammals, we crossed the viable rescued ADAR1-editing-deficient animals (Adar1E861A/E861AIfih1-/-) with rescued ADAR2-deficient (Adarb1-/-Gria2R/R) animals. Unexpectedly, the global absence of editing was well tolerated. Adar1E861A/E861AIfih1-/-Adarb1-/-Gria2R/R were recovered at Mendelian ratios and age normally. Detailed transcriptome analysis demonstrated that editing was absent in the brains of the compound mutants and that ADAR1 and ADAR2 have similar editing site preferences and patterns.
Conclusions: We conclude that ADAR1 and ADAR2 are non-redundant and do not compensate for each other's essential functions in vivo. Physiologically essential A-to-I editing comprises a small subset of the editome, and the majority of editing is dispensable for mammalian homeostasis. Moreover, in vivo biologically essential protein recoding mediated by A-to-I editing is an exception in mammals.
Keywords: A-to-I editing; ADAR1; ADAR2; Epitranscriptome; RNA editing; RNA modification.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
