

Understanding Long Noncoding RNA and Chromatin Interactions: What We Know So Far
- PMID: 31817041
- PMCID: PMC6958424
- DOI: 10.3390/ncrna5040054
Understanding Long Noncoding RNA and Chromatin Interactions: What We Know So Far
Abstract
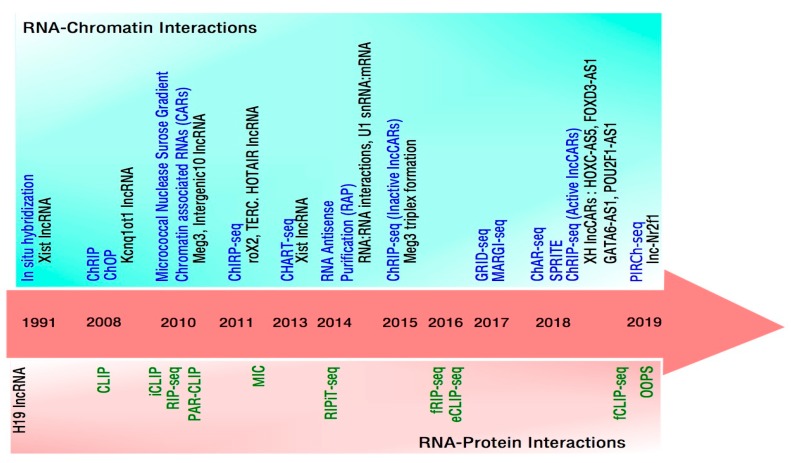
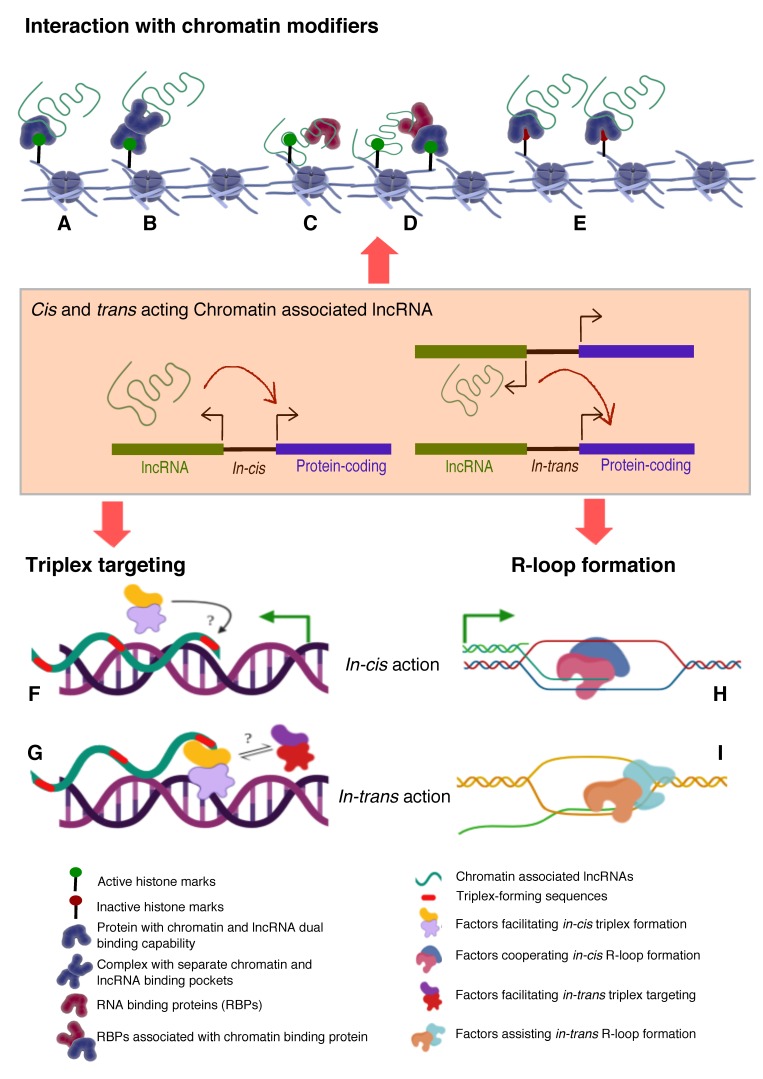
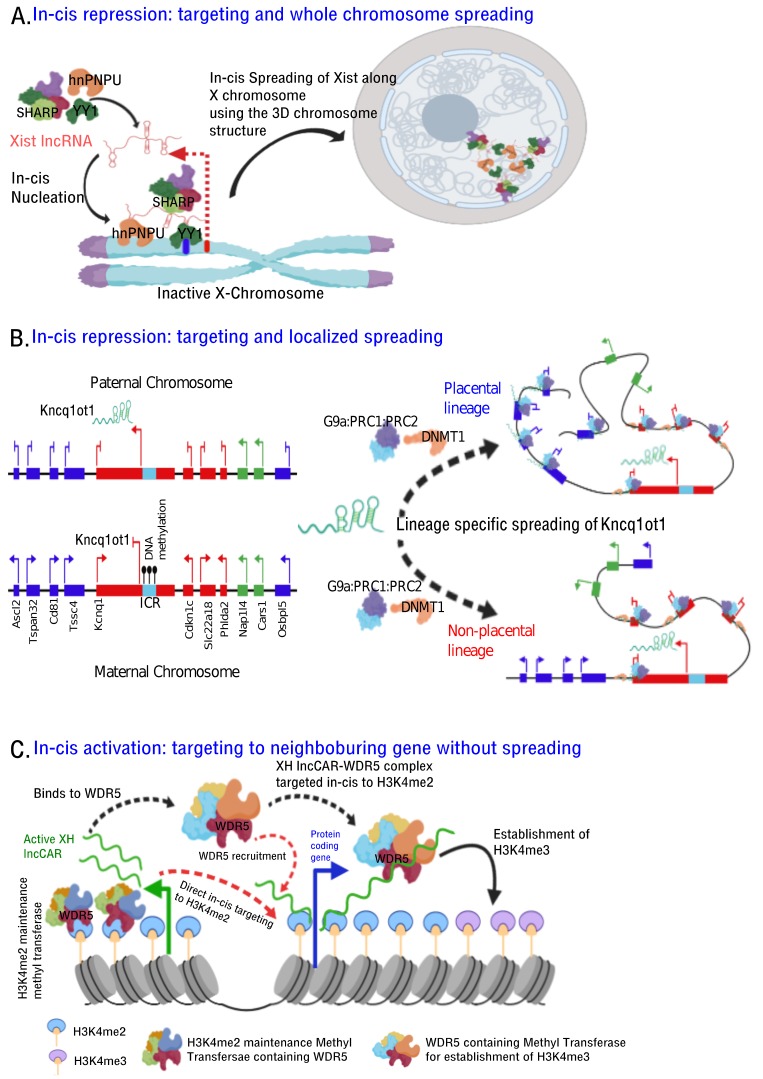
With the evolution of technologies that deal with global detection of RNAs to probing of lncRNA-chromatin interactions and lncRNA-chromatin structure regulation, we have been updated with a comprehensive repertoire of chromatin interacting lncRNAs, their genome-wide chromatin binding regions and mode of action. Evidence from these new technologies emphasize that chromatin targeting of lncRNAs is a prominent mechanism and that these chromatin targeted lncRNAs exert their functionality by fine tuning chromatin architecture resulting in an altered transcriptional readout. Currently, there are no unifying principles that define chromatin association of lncRNAs, however, evidence from a few chromatin-associated lncRNAs show presence of a short common sequence for chromatin targeting. In this article, we review how technological advancements contributed in characterizing chromatin associated lncRNAs, and discuss the potential mechanisms by which chromatin associated lncRNAs execute their functions.
Keywords: RNA-chromatin interactions; chromatin; chromatin RNA; gene regulation; lncrna; long noncoding RNA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
