

Distinct patterns of complex rearrangements and a mutational signature of microhomeology are frequently observed in PLP1 copy number gain structural variants
- PMID: 31818324
- PMCID: PMC6902434
- DOI: 10.1186/s13073-019-0676-0
Distinct patterns of complex rearrangements and a mutational signature of microhomeology are frequently observed in PLP1 copy number gain structural variants
Abstract
Background: We investigated the features of the genomic rearrangements in a cohort of 50 male individuals with proteolipid protein 1 (PLP1) copy number gain events who were ascertained with Pelizaeus-Merzbacher disease (PMD; MIM: 312080). We then compared our new data to previous structural variant mutagenesis studies involving the Xq22 region of the human genome. The aggregate data from 159 sequenced join-points (discontinuous sequences in the reference genome that are joined during the rearrangement process) were studied. Analysis of these data from 150 individuals enabled the spectrum and relative distribution of the underlying genomic mutational signatures to be delineated.
Methods: Genomic rearrangements in PMD individuals with PLP1 copy number gain events were investigated by high-density customized array or clinical chromosomal microarray analysis and breakpoint junction sequence analysis.
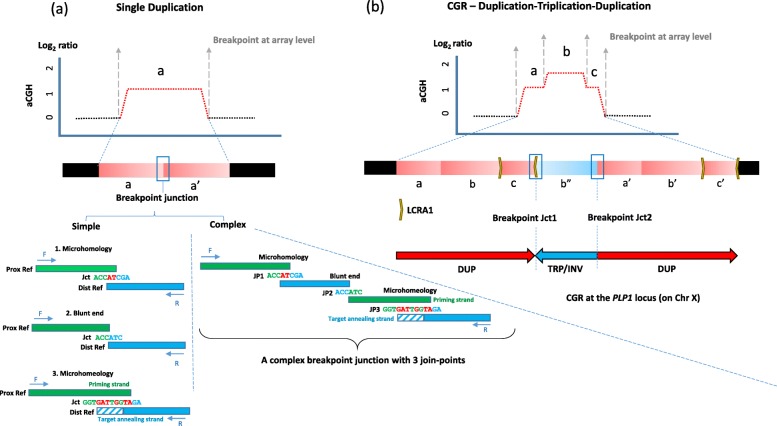
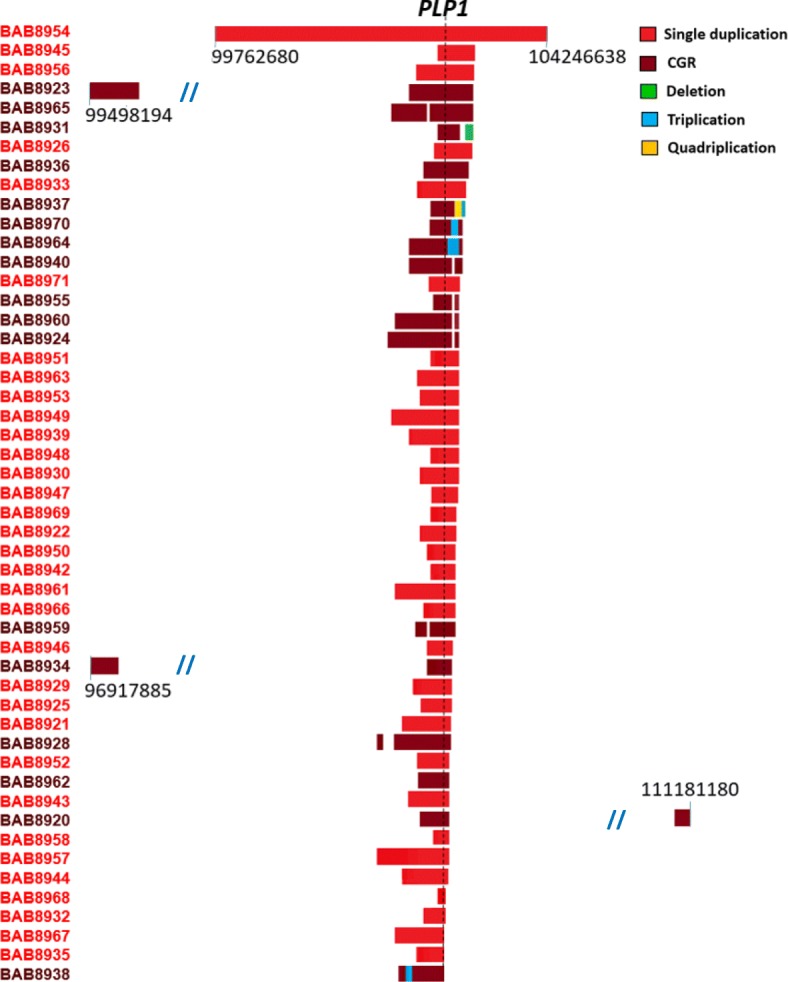
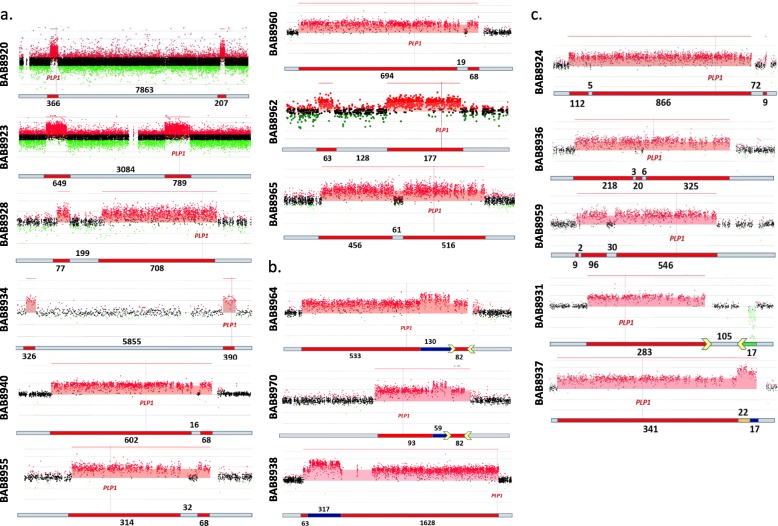
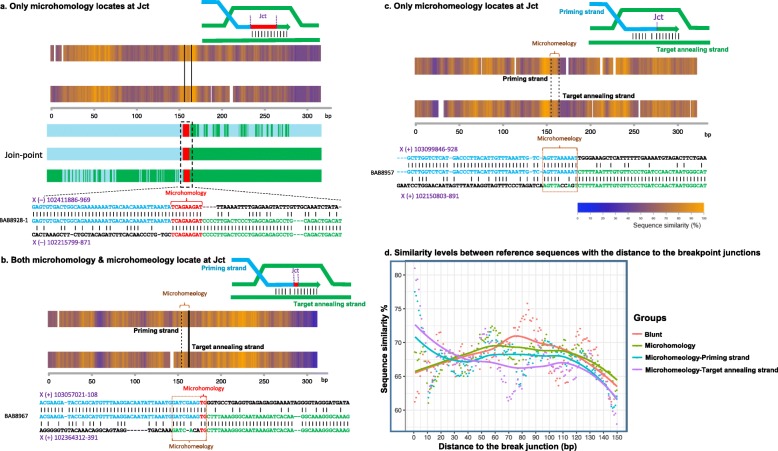
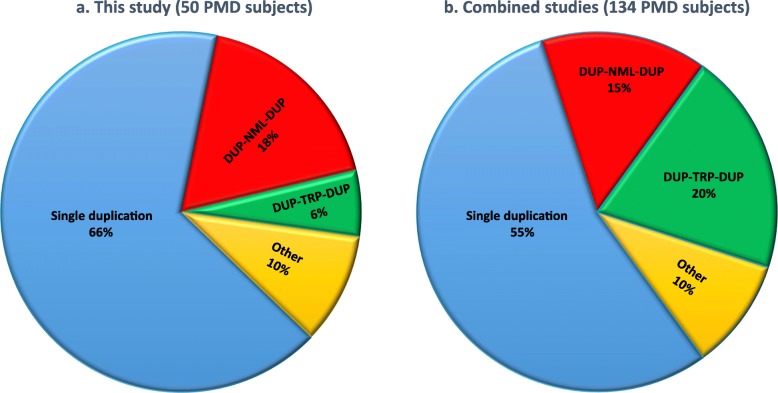
Results: High-density customized array showed that the majority of cases (33/50; ~ 66%) present with single duplications, although complex genomic rearrangements (CGRs) are also frequent (17/50; ~ 34%). Breakpoint mapping to nucleotide resolution revealed further previously unknown structural and sequence complexities, even in single duplications. Meta-analysis of all studied rearrangements that occur at the PLP1 locus showed that single duplications were found in ~ 54% of individuals and that, among all CGR cases, triplication flanked by duplications is the most frequent CGR array CGH pattern observed. Importantly, in ~ 32% of join-points, there is evidence for a mutational signature of microhomeology (highly similar yet imperfect sequence matches).
Conclusions: These data reveal a high frequency of CGRs at the PLP1 locus and support the assertion that replication-based mechanisms are prominent contributors to the formation of CGRs at Xq22. We propose that microhomeology can facilitate template switching, by stabilizing strand annealing of the primer using W-C base complementarity, and is a mutational signature for replicative repair.
Keywords: BIR; Duplication; Genome instability; Genomic rearrangements; HR; LCR; MMBIR; Microhomeology; PMD; RBM.
Conflict of interest statement
J.R.L. has stock ownership in 23andMe, is a paid consultant for Regeneron Pharmaceuticals, and is a co-inventor on multiple US and European patents related to molecular diagnostics for inherited neuropathies, eye diseases, and bacterial genomic fingerprinting. The Department of Molecular and Human Genetics at Baylor College of Medicine derives revenue from the chromosomal microarray analysis (CMA) and clinical exome sequencing offered in the Baylor Genetics Laboratory (BMGL:
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
