

Genetic and Epigenetic Control of Heart Development
- PMID: 31818853
- PMCID: PMC7280077
- DOI: 10.1101/cshperspect.a036756
Genetic and Epigenetic Control of Heart Development
Abstract
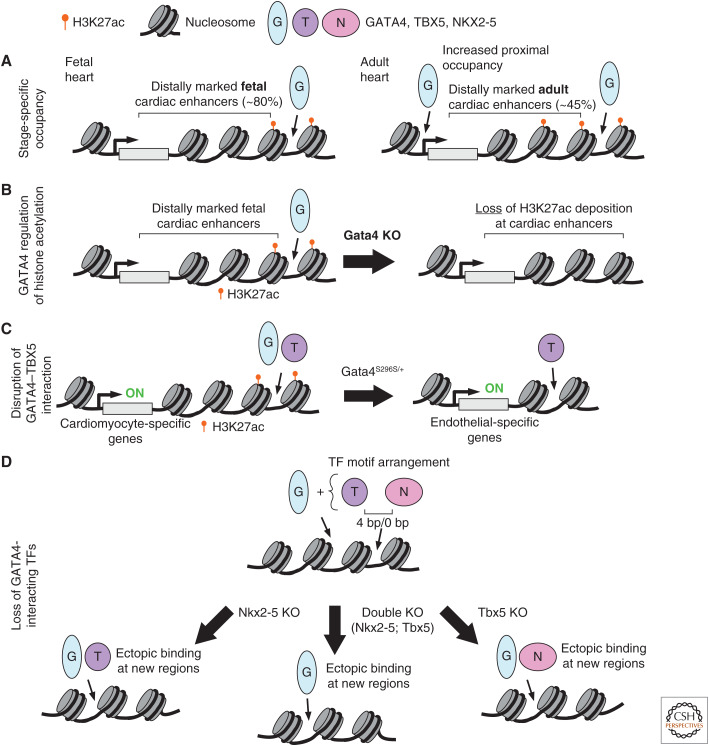
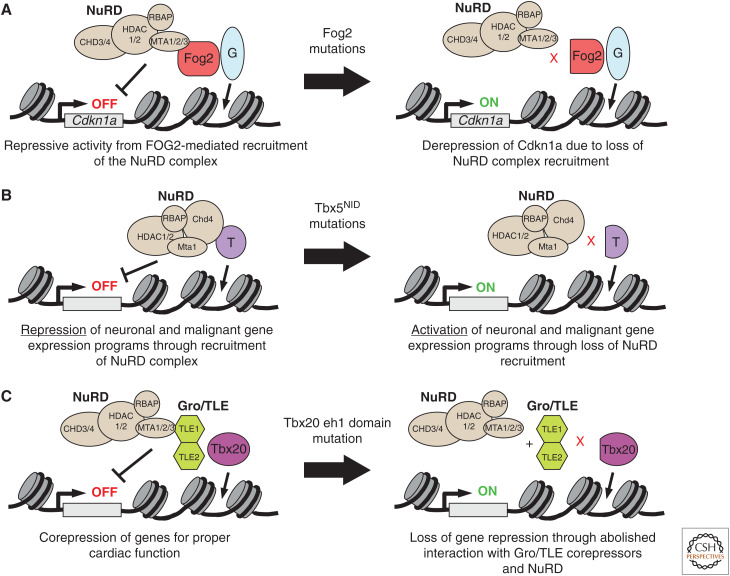
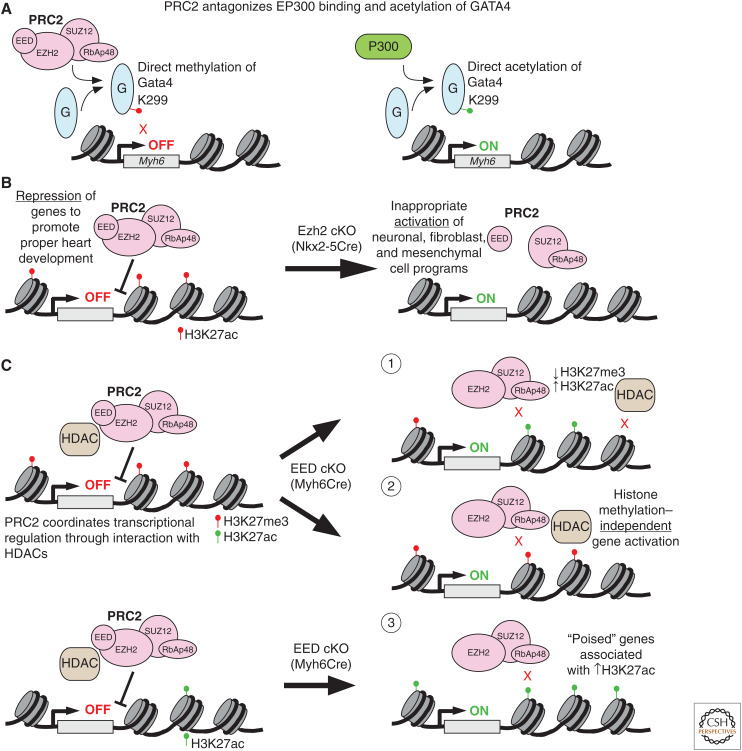
A transcriptional program implemented by transcription factors and epigenetic regulators governs cardiac development and disease. Mutations in these factors are important causes of congenital heart disease. Here, we review selected recent advances in our understanding of the transcriptional and epigenetic control of heart development, including determinants of cardiac transcription factor chromatin occupancy, the gene regulatory network that regulates atrial septation, the chromatin landscape and cardiac gene regulation, and the role of Brg/Brahma-associated factor (BAF), nucleosome remodeling and histone deacetylation (NuRD), and Polycomb epigenetic regulatory complexes in heart development.
Copyright © 2020 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources