

Dynamics of Candidatus Liberibacter asiaticus Movement and Sieve-Pore Plugging in Citrus Sink Cells
- PMID: 31818905
- PMCID: PMC6997701
- DOI: 10.1104/pp.19.01391
Dynamics of Candidatus Liberibacter asiaticus Movement and Sieve-Pore Plugging in Citrus Sink Cells
Abstract
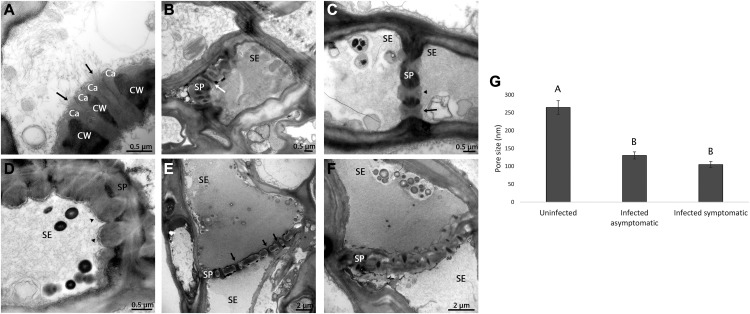
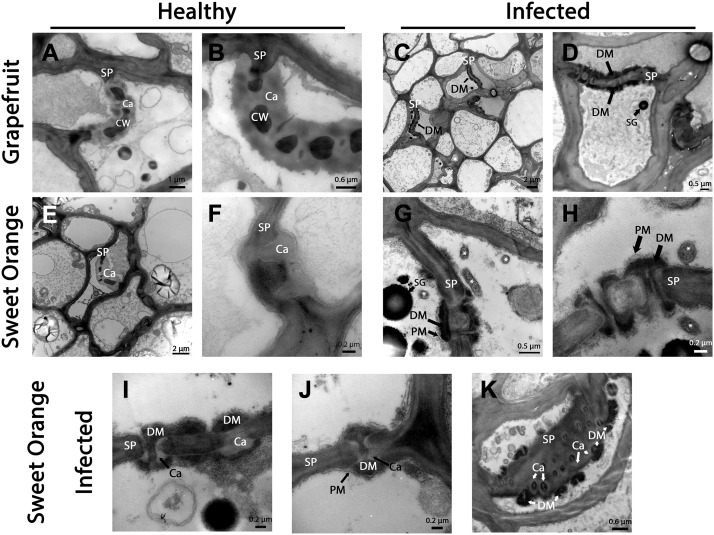
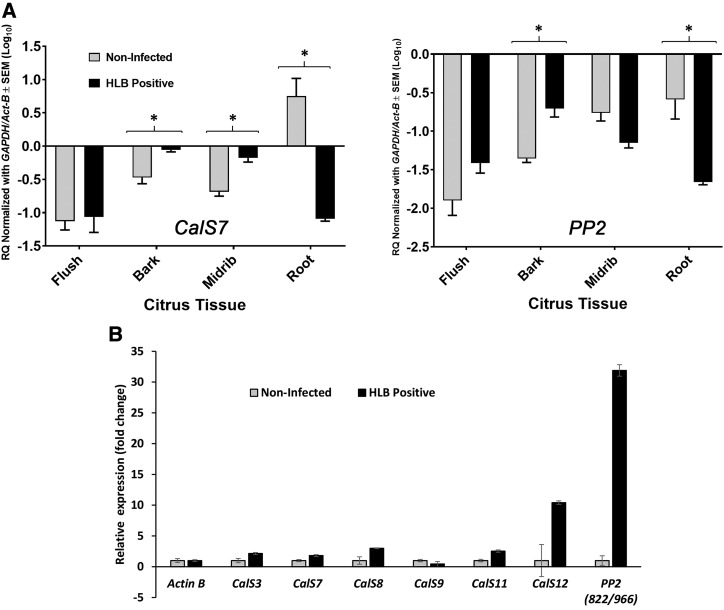
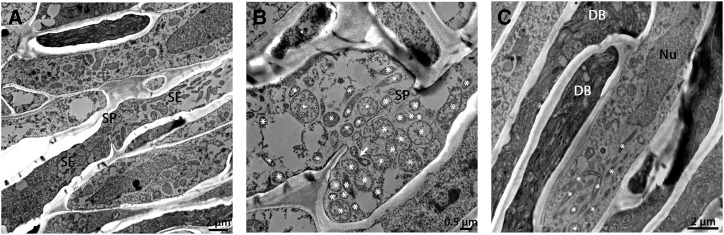
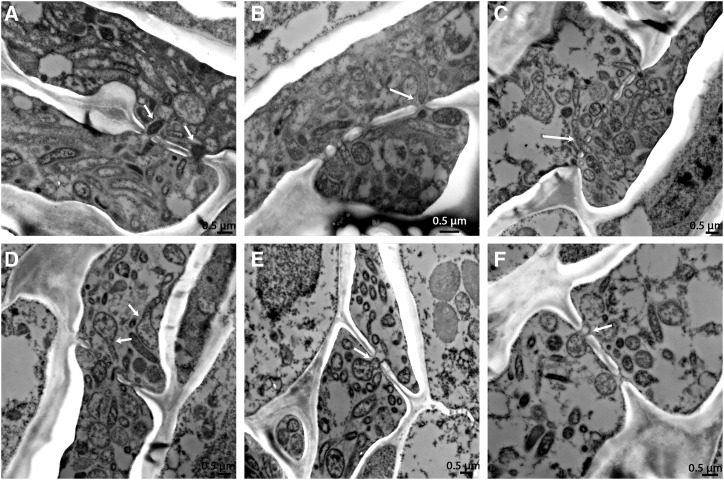
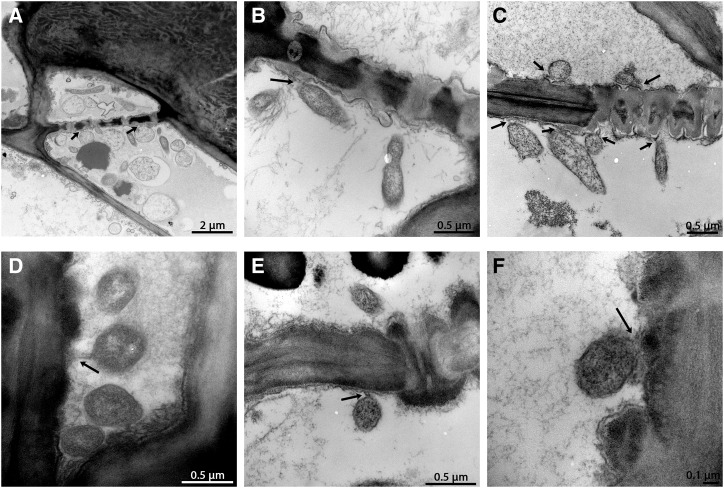
Citrus greening or Huanglongbing (HLB) is caused by the phloem-limited intracellular Gram-negative bacterium Candidatus Liberibacter asiaticus (CLas). HLB-infected citrus phloem cells undergo structural modifications that include cell wall thickening, callose and phloem protein induction, and cellular plugging. However, very little is known about the intracellular mechanisms that take place during CLas cell-to-cell movement. Here, we show that CLas movement through phloem pores of sweet orange (Citrus sinensis) and grapefruit (Citrus paradisi) is carried out by the elongated form of the bacteria. The round form of CLas is too large to move, but can change its morphology to enable its movement. CLas cells adhere to the plasma membrane of the phloem cells specifically adjacent to the sieve pores. Remarkably, CLas was present in both mature sieve element cells and nucleated nonsieve element cells. The sieve plate plugging structures of host plants were shown to have different composition in different citrus tissues. Callose deposition was the main plugging mechanism in the HLB-infected flush, where it reduced the open space of the pores. In the roots, pores were surrounded by dark extracellular material, with very little accumulation of callose. The expression of CALLOSE SYNTHASE7 and PHLOEM PROTEIN2 genes was upregulated in the shoots, but downregulated in root tissues. In seed coats, no phloem occlusion was observed, and CLas accumulated to high levels. Our results provide insight into the cellular mechanisms of Gram-negative bacterial cell-to-cell movement in plant phloem.
© 2020 American Society of Plant Biologists. All Rights Reserved.
Figures
Similar articles
-
Callose deposition in the phloem plasmodesmata and inhibition of phloem transport in citrus leaves infected with "Candidatus Liberibacter asiaticus".Protoplasma. 2012 Jul;249(3):687-97. doi: 10.1007/s00709-011-0312-3. Epub 2011 Aug 28. Protoplasma. 2012. PMID: 21874517
-
Response of sweet orange (Citrus sinensis) to 'Candidatus Liberibacter asiaticus' infection: microscopy and microarray analyses.Phytopathology. 2009 Jan;99(1):50-7. doi: 10.1094/PHYTO-99-1-0050. Phytopathology. 2009. PMID: 19055434
-
Physiologic, Anatomic, and Gene Expression Changes in Citrus sunki, Poncirus trifoliata, and Their Hybrids After 'Candidatus Liberibacter asiaticus' Infection.Phytopathology. 2017 May;107(5):590-599. doi: 10.1094/PHYTO-02-16-0077-R. Epub 2017 Mar 24. Phytopathology. 2017. PMID: 28068188
-
Progress and Obstacles in Culturing 'Candidatus Liberibacter asiaticus', the Bacterium Associated with Huanglongbing.Phytopathology. 2019 Jul;109(7):1092-1101. doi: 10.1094/PHYTO-02-19-0051-RVW. Epub 2019 Jun 3. Phytopathology. 2019. PMID: 30998129 Review.
-
Molecular signatures between citrus and Candidatus Liberibacter asiaticus.PLoS Pathog. 2021 Dec 9;17(12):e1010071. doi: 10.1371/journal.ppat.1010071. eCollection 2021 Dec. PLoS Pathog. 2021. PMID: 34882744 Free PMC article. Review.
Cited by
-
Proteomic and bioinformatic analyses of proteins in the outer membrane and extracellular compartments and outer membrane vesicles of Candidatus Liberibacter species.Front Microbiol. 2022 Sep 26;13:977710. doi: 10.3389/fmicb.2022.977710. eCollection 2022. Front Microbiol. 2022. PMID: 36225379 Free PMC article.
-
Better tolerance to Huanglongbing is conferred by tetraploid Swingle citrumelo rootstock and is influenced by the ploidy of the scion.Front Plant Sci. 2022 Nov 3;13:1030862. doi: 10.3389/fpls.2022.1030862. eCollection 2022. Front Plant Sci. 2022. PMID: 36407590 Free PMC article.
-
Plasmodesmata-Involved Battle Against Pathogens and Potential Strategies for Strengthening Hosts.Front Plant Sci. 2021 Jun 3;12:644870. doi: 10.3389/fpls.2021.644870. eCollection 2021. Front Plant Sci. 2021. PMID: 34149749 Free PMC article. Review.
-
A serralysin-like protein of Candidatus Liberibacter asiaticus modulates components of the bacterial extracellular matrix.Front Microbiol. 2022 Oct 19;13:1006962. doi: 10.3389/fmicb.2022.1006962. eCollection 2022. Front Microbiol. 2022. PMID: 36338045 Free PMC article.
-
A Significantly High Abundance of "Candidatus Liberibacter asiaticus" in Citrus Fruit Pith: in planta Transcriptome and Anatomical Analyses.Front Microbiol. 2021 Jun 11;12:681251. doi: 10.3389/fmicb.2021.681251. eCollection 2021. Front Microbiol. 2021. PMID: 34177866 Free PMC article.
References
-
- Achor DS, Etxeberria E, Wang N, Folimonova SY, Chung KR, Albrigo LB (2010) Sequence of anatomical symptom observations in citrus affected with Huanglongbing disease. Plant Pathol J 9: 56–64
-
- Aloni R, Raviv A, Peterson CA (1991) The role of auxin in the removal of dormancy callose and resumption of phloem activity in Vitis vinifera. Canadian J Botany-Revue Canadienne De Botanique 69: 1825–1832
-
- Behnke HD, Sjolund RD (1990) Sieve Elements. Springer, Berlin, Heidelberg, New York
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
