

Transcriptomic analysis of human primary breast cancer identifies fatty acid oxidation as a target for metformin
- PMID: 31819193
- PMCID: PMC6986920
- DOI: 10.1038/s41416-019-0665-5
Transcriptomic analysis of human primary breast cancer identifies fatty acid oxidation as a target for metformin
Abstract
Background: Epidemiological studies suggest that metformin may reduce the incidence of cancer in patients with diabetes and multiple late phase clinical trials assessing the potential of repurposing this drug are underway. Transcriptomic profiling of tumour samples is an excellent tool to understand drug bioactivity, identify candidate biomarkers and assess for mechanisms of resistance to therapy.
Methods: Thirty-six patients with untreated primary breast cancer were recruited to a window study and transcriptomic profiling of tumour samples carried out before and after metformin treatment.
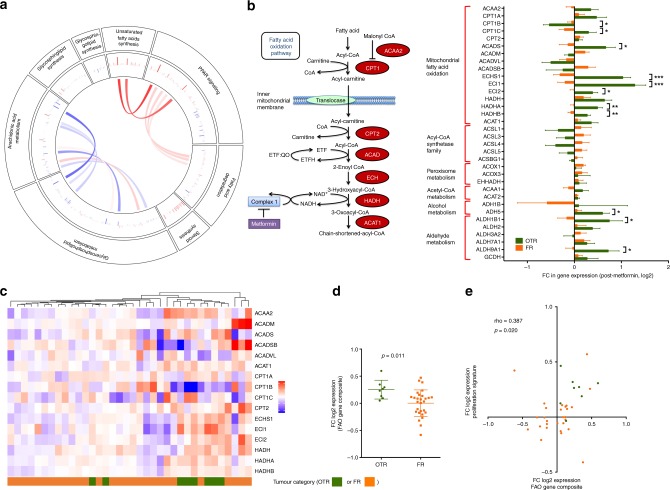
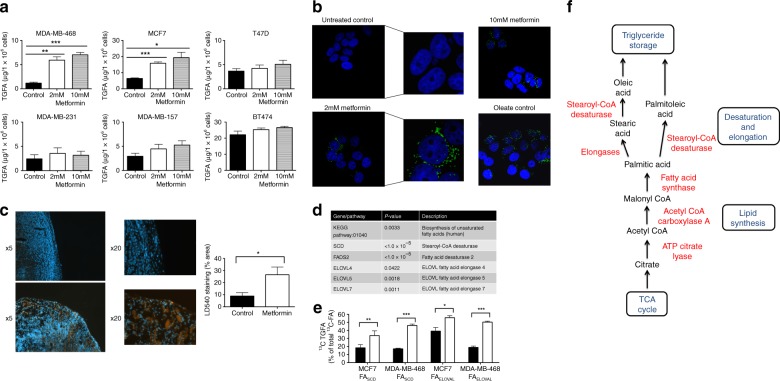
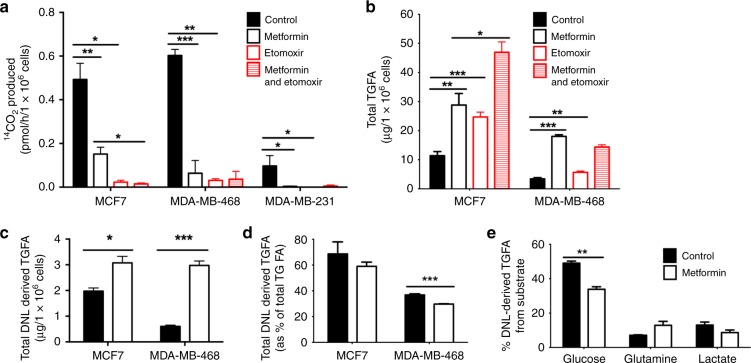
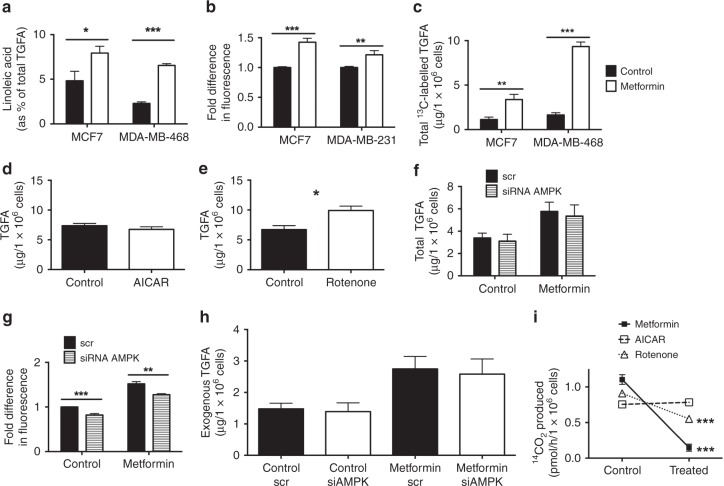
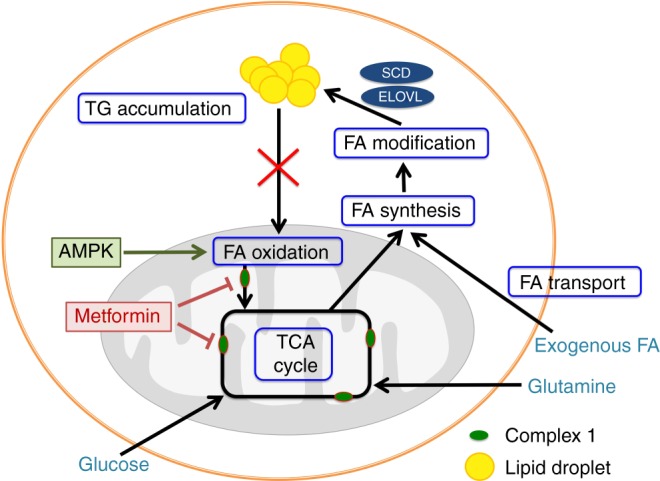
Results: Multiple genes that regulate fatty acid oxidation were upregulated at the transcriptomic level and there was a differential change in expression between two previously identified cohorts of patients with distinct metabolic responses. Increase in expression of a mitochondrial fatty oxidation gene composite signature correlated with change in a proliferation gene signature. In vitro assays showed that, in contrast to previous studies in models of normal cells, metformin reduces fatty acid oxidation with a subsequent accumulation of intracellular triglyceride, independent of AMPK activation.
Conclusions: We propose that metformin at clinical doses targets fatty acid oxidation in cancer cells with implications for patient selection and drug combinations.
Clinical trial registration: NCT01266486.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Lord Simon R., Cheng Wei-Chen, Liu Dan, Gaude Edoardo, Haider Syed, Metcalf Tom, Patel Neel, Teoh Eugene J., Gleeson Fergus, Bradley Kevin, Wigfield Simon, Zois Christos, McGowan Daniel R., Ah-See Mei-Lin, Thompson Alastair M., Sharma Anand, Bidaut Luc, Pollak Michael, Roy Pankaj G., Karpe Fredrik, James Tim, English Ruth, Adams Rosie F., Campo Leticia, Ayers Lisa, Snell Cameron, Roxanis Ioannis, Frezza Christian, Fenwick John D., Buffa Francesca M., Harris Adrian L. Integrated Pharmacodynamic Analysis Identifies Two Metabolic Adaption Pathways to Metformin in Breast Cancer. Cell Metabolism. 2018;28(5):679-688.e4. doi: 10.1016/j.cmet.2018.08.021. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
