

Reactivation of Epstein-Barr virus by a dual-responsive fluorescent EBNA1-targeting agent with Zn2+-chelating function
- PMID: 31822610
- PMCID: PMC6936348
- DOI: 10.1073/pnas.1915372116
Reactivation of Epstein-Barr virus by a dual-responsive fluorescent EBNA1-targeting agent with Zn2+-chelating function
Erratum in
-
Correction for Jiang et al., Reactivation of Epstein-Barr virus by a dual-responsive fluorescent EBNA1-targeting agent with Zn2+-chelating function.Proc Natl Acad Sci U S A. 2020 Mar 10;117(10):5542. doi: 10.1073/pnas.2002334117. Epub 2020 Mar 2. Proc Natl Acad Sci U S A. 2020. PMID: 32123071 Free PMC article. No abstract available.
Abstract
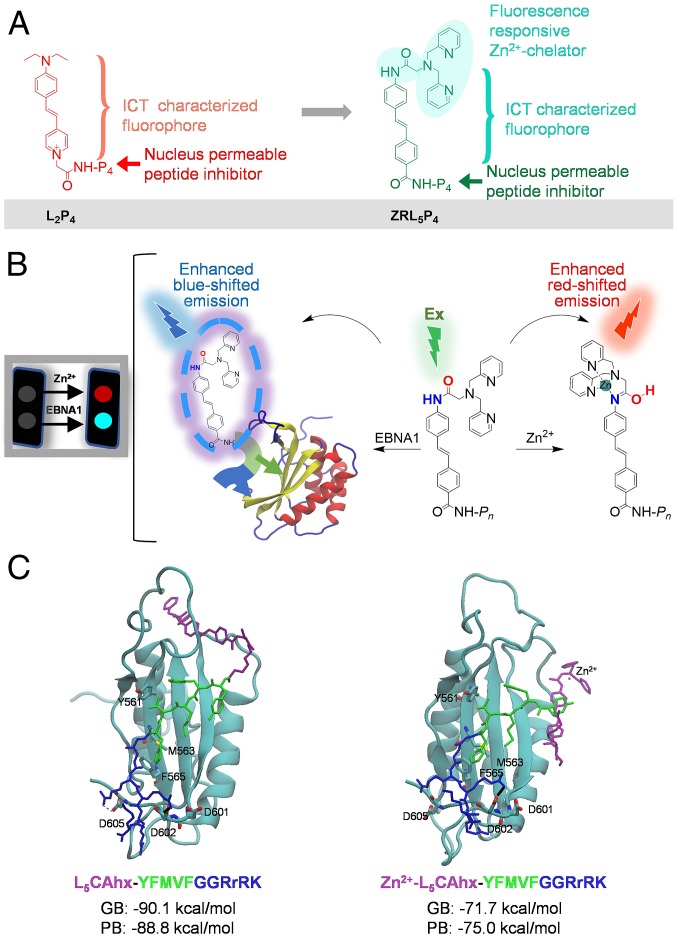
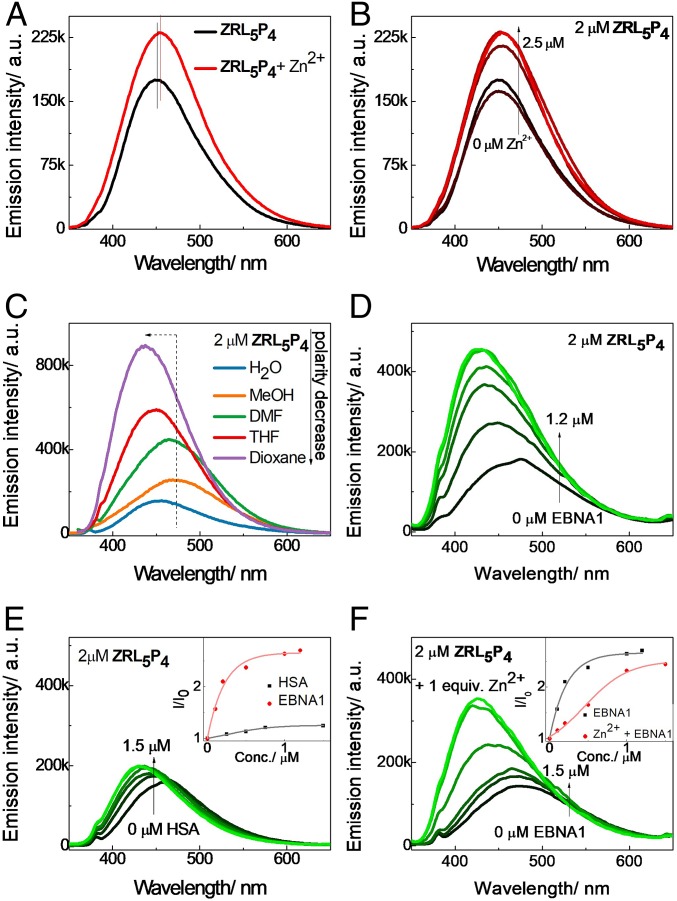
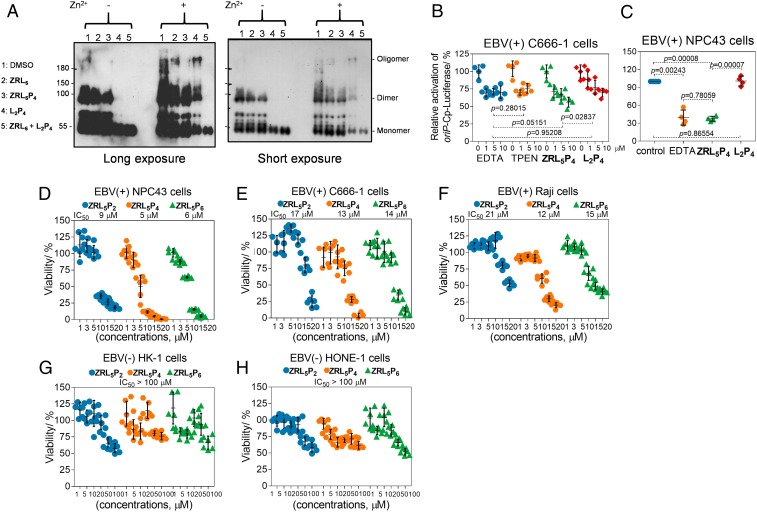
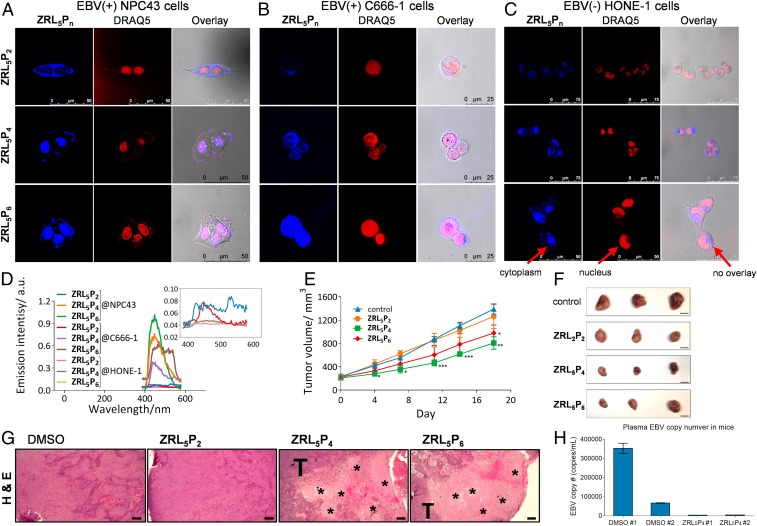
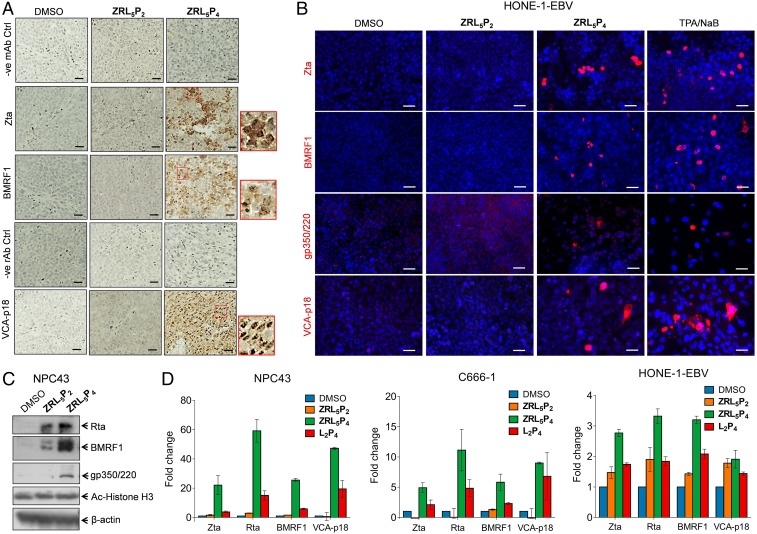
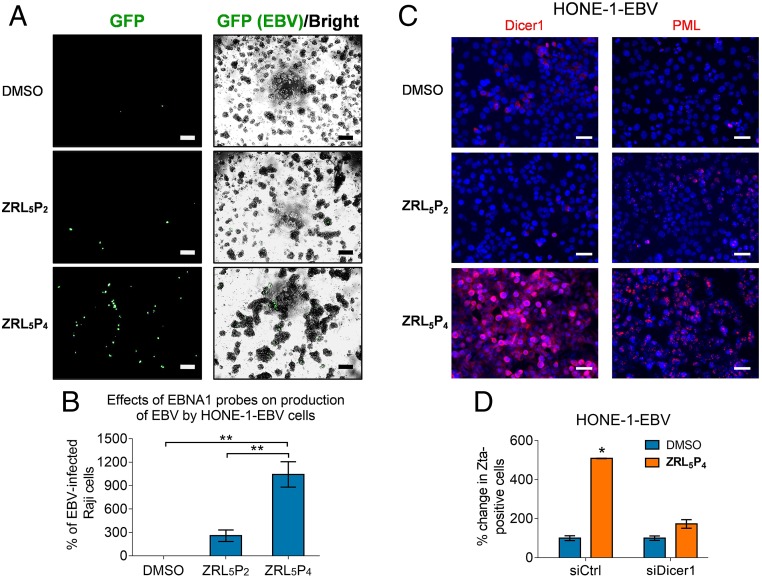
Epstein-Barr nuclear antigen 1 (EBNA1) plays a vital role in the maintenance of the viral genome and is the only viral protein expressed in nearly all forms of Epstein-Barr virus (EBV) latency and EBV-associated diseases, including numerous cancer types. To our knowledge, no specific agent against EBV genes or proteins has been established to target EBV lytic reactivation. Here we report an EBNA1- and Zn2+-responsive probe (ZRL5P4) which alone could reactivate the EBV lytic cycle through specific disruption of EBNA1. We have utilized the Zn2+ chelator to further interfere with the higher order of EBNA1 self-association. The bioprobe ZRL5P4 can respond independently to its interactions with Zn2+ and EBNA1 with different fluorescence changes. It can selectively enter the nuclei of EBV-positive cells and disrupt the oligomerization and oriP-enhanced transactivation of EBNA1. ZRL5P4 can also specifically enhance Dicer1 and PML expression, molecular events which had been reported to occur after the depletion of EBNA1 expression. Importantly, we found that treatment with ZRL5P4 alone could reactivate EBV lytic induction by expressing the early and late EBV lytic genes/proteins. Lytic induction is likely mediated by disruption of EBNA1 oligomerization and the subsequent change of Dicer1 expression. Our probe ZRL5P4 is an EBV protein-specific agent that potently reactivates EBV from latency, leading to the shrinkage of EBV-positive tumors, and our study also suggests the association of EBNA1 oligomerization with the maintenance of EBV latency.
Keywords: EBNA1-targeting agent; EBV-specific lytic inducer; dual-responsive fluorescent EBV probe.
Conflict of interest statement
Competing interest statement: K.-L.W., N.K.M., and L.J. are listed as inventors on a filed US nonprovisional patent titled “Zinc-binder based EBNA1-specific compounds” with the application number 16/249,987.
Figures
References
-
- Cohen J. I., Epstein-Barr virus infection. N. Engl. J. Med. 343, 481–492 (2000). - PubMed
-
- Novalić Z., Rossen T. M. v., Greijer A. E., Middeldorp J. M., Agents and approaches for lytic induction therapy of Epstein-Barr virus associated malignancies. Med. Chem. 6, 449–466 (2016).
LinkOut - more resources
Full Text Sources
Other Literature Sources
