

Spectroscopic and biochemical insight into an electron-bifurcating [FeFe] hydrogenase
- PMID: 31823008
- PMCID: PMC7064455
- DOI: 10.1007/s00775-019-01747-1
Spectroscopic and biochemical insight into an electron-bifurcating [FeFe] hydrogenase
Abstract
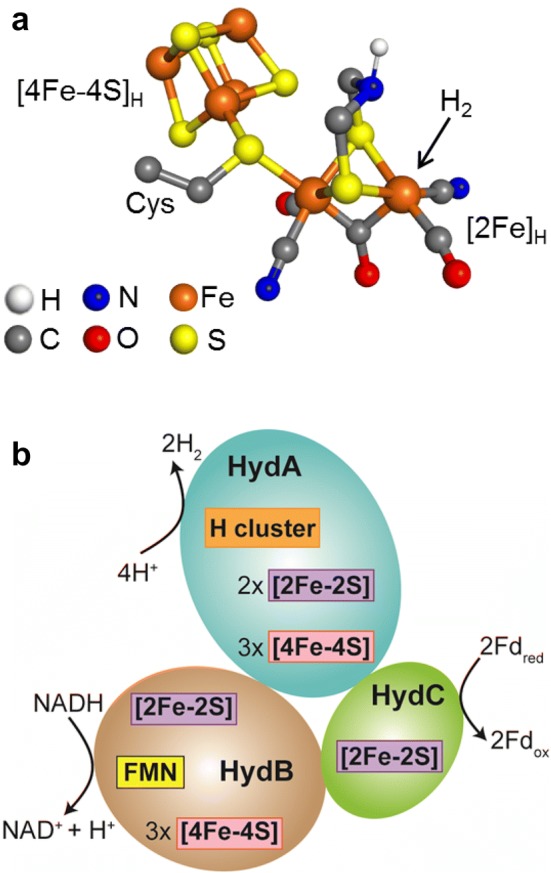
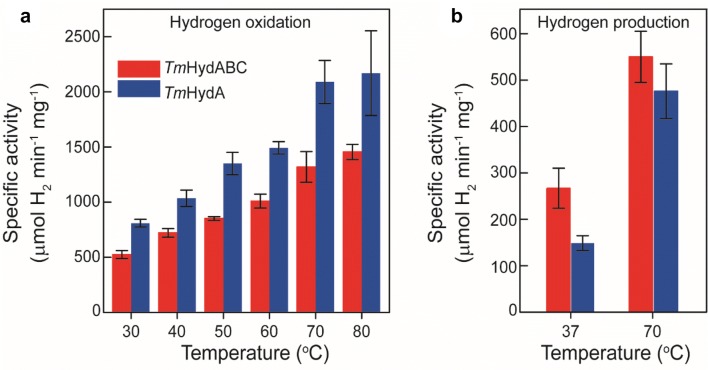
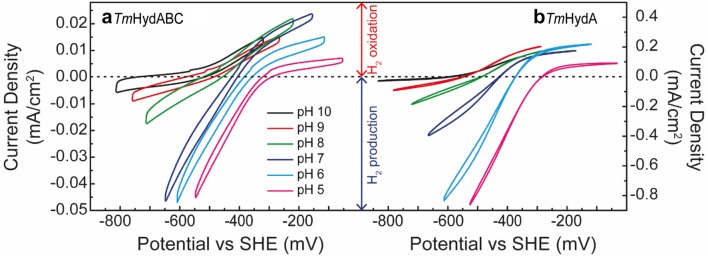
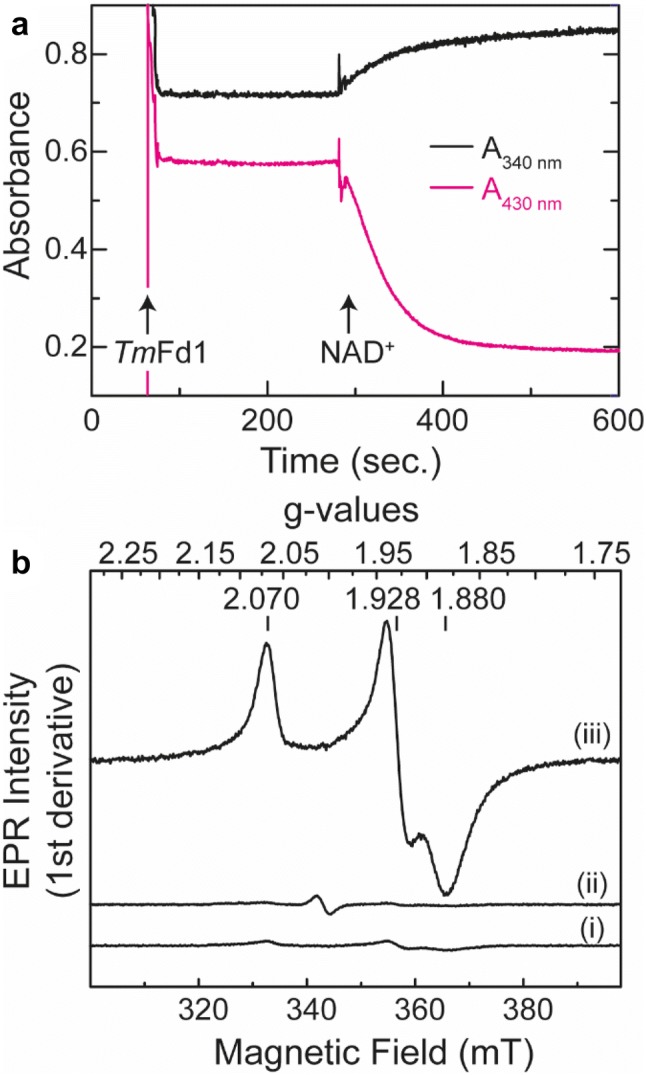
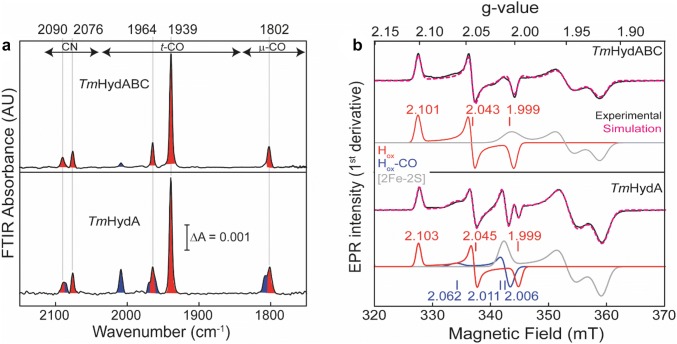
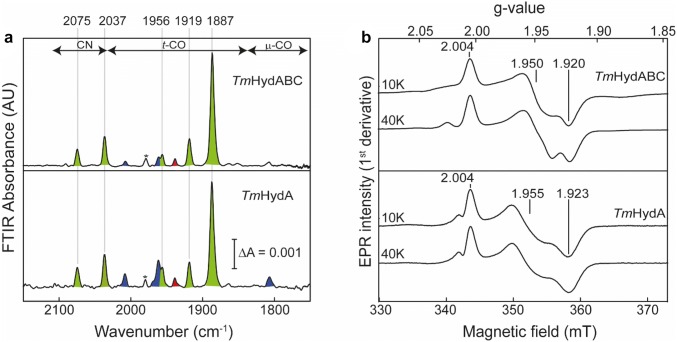
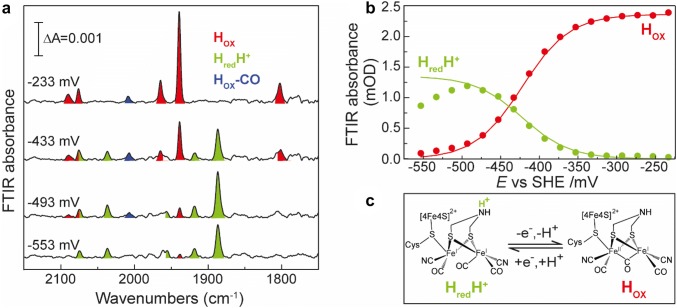
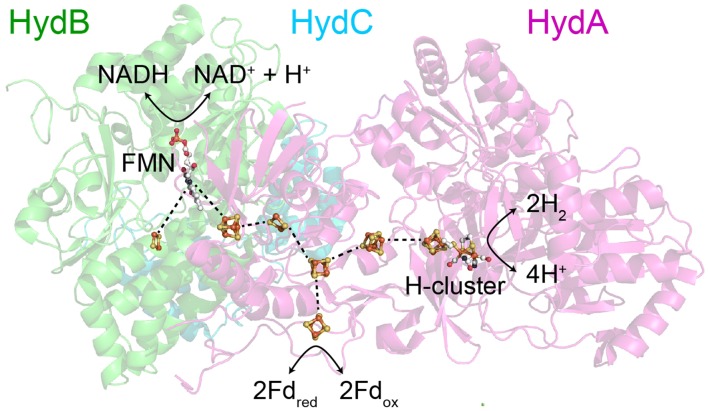
The heterotrimeric electron-bifurcating [FeFe] hydrogenase (HydABC) from Thermotoga maritima (Tm) couples the endergonic reduction of protons (H+) by dihydronicotinamide adenine dinucleotide (NADH) (∆G0 ≈ 18 kJ mol-1) to the exergonic reduction of H+ by reduced ferredoxin (Fdred) (∆G0 ≈ - 16 kJ mol-1). The specific mechanism by which HydABC functions is not understood. In the current study, we describe the biochemical and spectroscopic characterization of TmHydABC recombinantly produced in Escherichia coli and artificially maturated with a synthetic diiron cofactor. We found that TmHydABC catalyzed the hydrogen (H2)-dependent reduction of nicotinamide adenine dinucleotide (NAD+) in the presence of oxidized ferredoxin (Fdox) at a rate of ≈17 μmol NADH min-1 mg-1. Our data suggest that only one flavin is present in the enzyme and is not likely to be the site of electron bifurcation. FTIR and EPR spectroscopy, as well as FTIR spectroelectrochemistry, demonstrated that the active site for H2 conversion, the H-cluster, in TmHydABC behaves essentially the same as in prototypical [FeFe] hydrogenases, and is most likely also not the site of electron bifurcation. The implications of these results are discussed with respect to the current hypotheses on the electron bifurcation mechanism of [FeFe] hydrogenases. Overall, the results provide insight into the electron-bifurcating mechanism and present a well-defined system for further investigations of this fascinating class of [FeFe] hydrogenases.
Keywords: Electrochemistry; Electron bifurcation; Ferredoxin; Spectroscopy; [FeFe] hydrogenase.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
