

p27 allosterically activates cyclin-dependent kinase 4 and antagonizes palbociclib inhibition
- PMID: 31831640
- PMCID: PMC7592119
- DOI: 10.1126/science.aaw2106
p27 allosterically activates cyclin-dependent kinase 4 and antagonizes palbociclib inhibition
Abstract
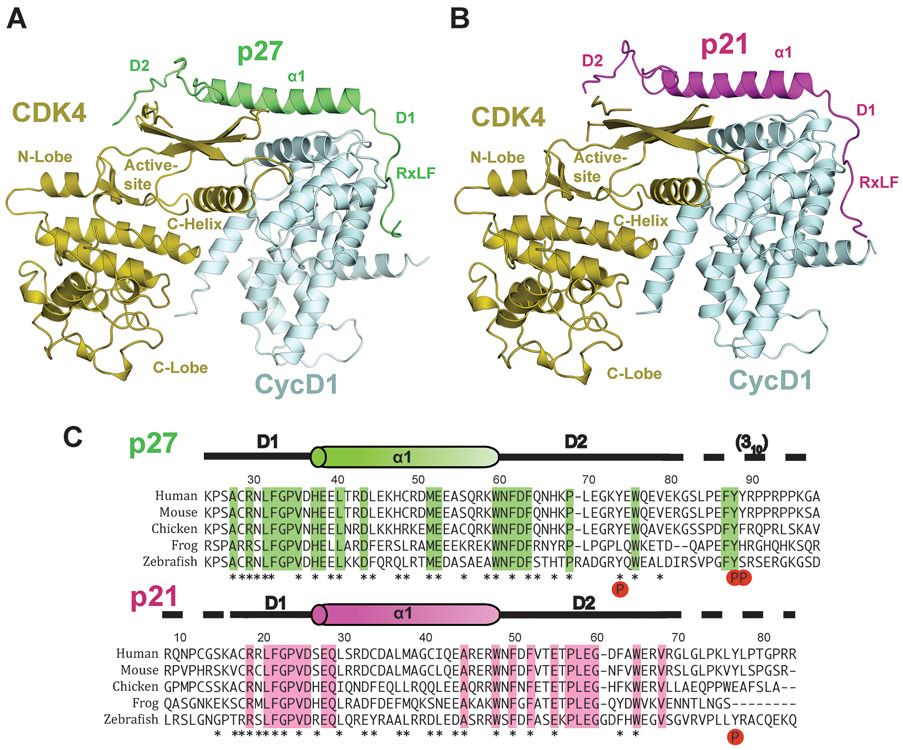
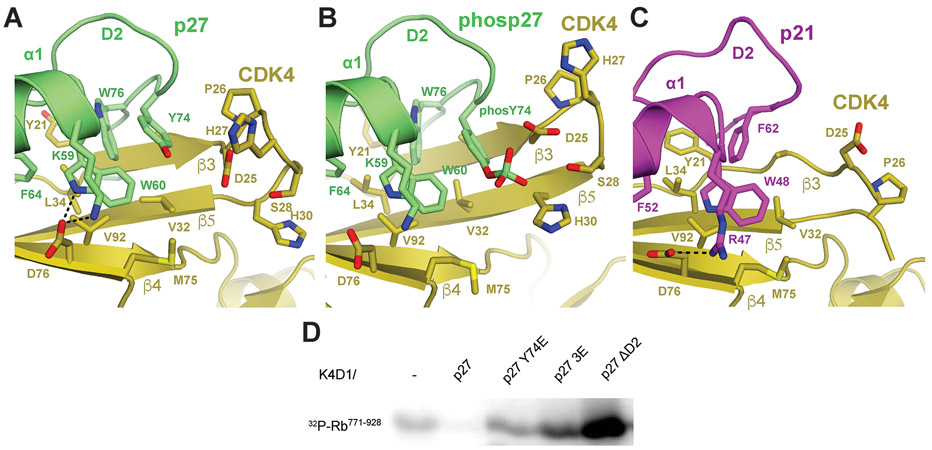
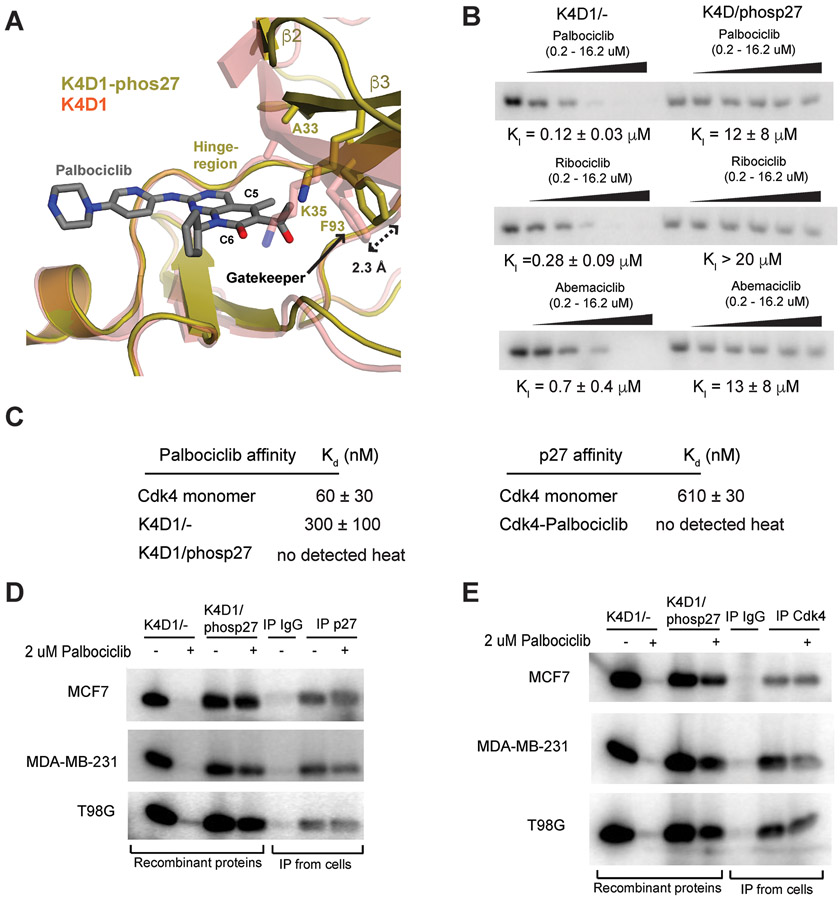
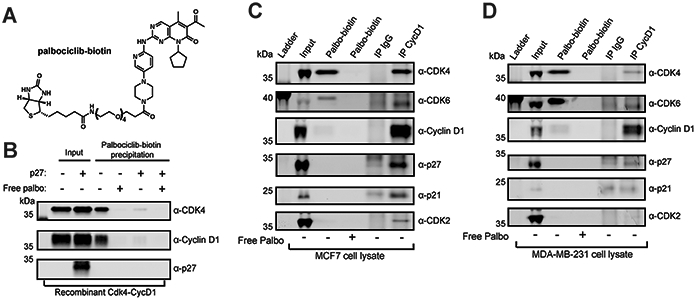
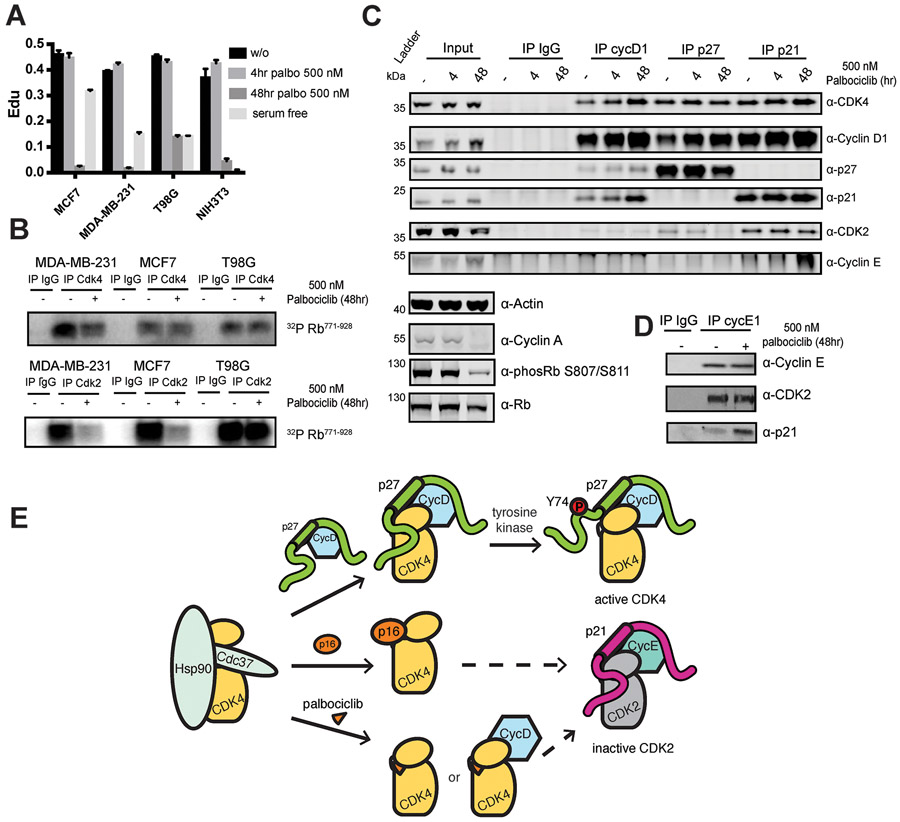
The p27 protein is a canonical negative regulator of cell proliferation and acts primarily by inhibiting cyclin-dependent kinases (CDKs). Under some circumstances, p27 is associated with active CDK4, but no mechanism for activation has been described. We found that p27, when phosphorylated by tyrosine kinases, allosterically activated CDK4 in complex with cyclin D1 (CDK4-CycD1). Structural and biochemical data revealed that binding of phosphorylated p27 (phosp27) to CDK4 altered the kinase adenosine triphosphate site to promote phosphorylation of the retinoblastoma tumor suppressor protein (Rb) and other substrates. Surprisingly, purified and endogenous phosp27-CDK4-CycD1 complexes were insensitive to the CDK4-targeting drug palbociclib. Palbociclib instead primarily targeted monomeric CDK4 and CDK6 (CDK4/6) in breast tumor cells. Our data characterize phosp27-CDK4-CycD1 as an active Rb kinase that is refractory to clinically relevant CDK4/6 inhibitors.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures



Comment in
-
Surprising regulation of cell cycle entry.Science. 2019 Dec 13;366(6471):1315-1316. doi: 10.1126/science.aaz4043. Science. 2019. PMID: 31831659 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
