

Strong suppression of gene conversion with increasing DNA double-strand break load delimited by 53BP1 and RAD52
- PMID: 31832684
- PMCID: PMC7038941
- DOI: 10.1093/nar/gkz1167
Strong suppression of gene conversion with increasing DNA double-strand break load delimited by 53BP1 and RAD52
Abstract
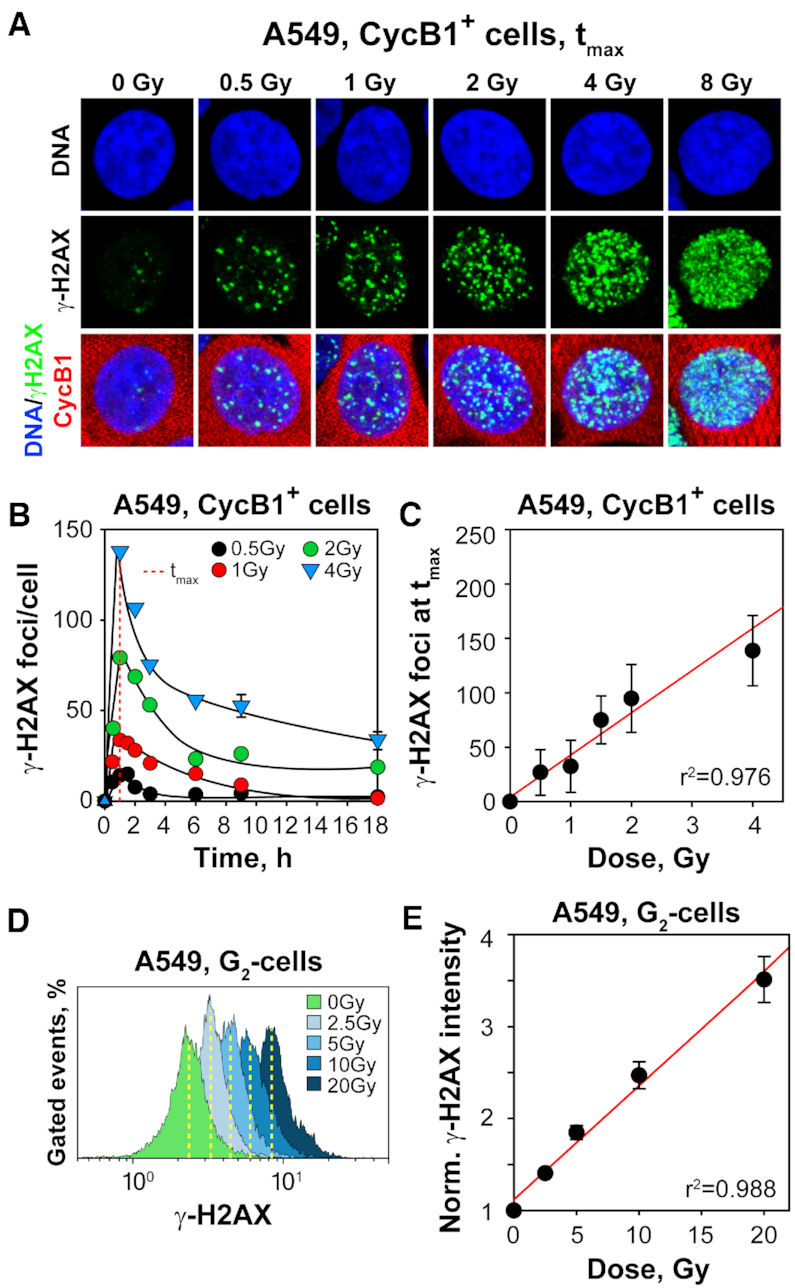
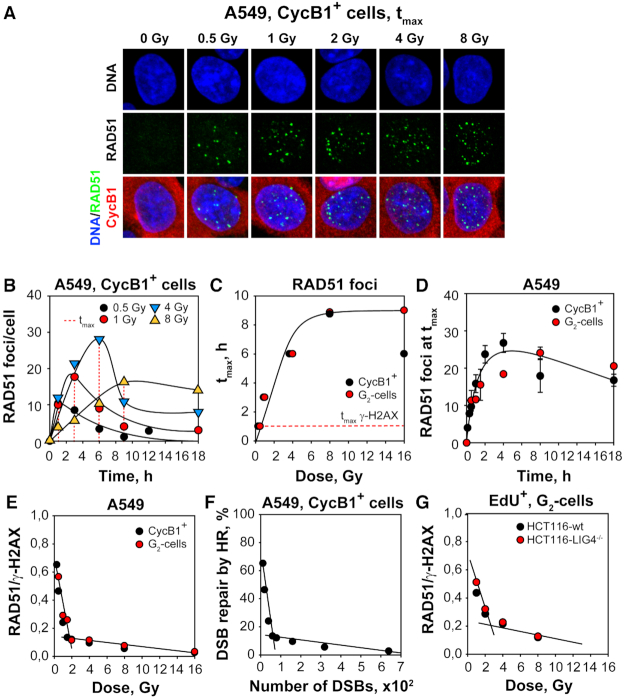
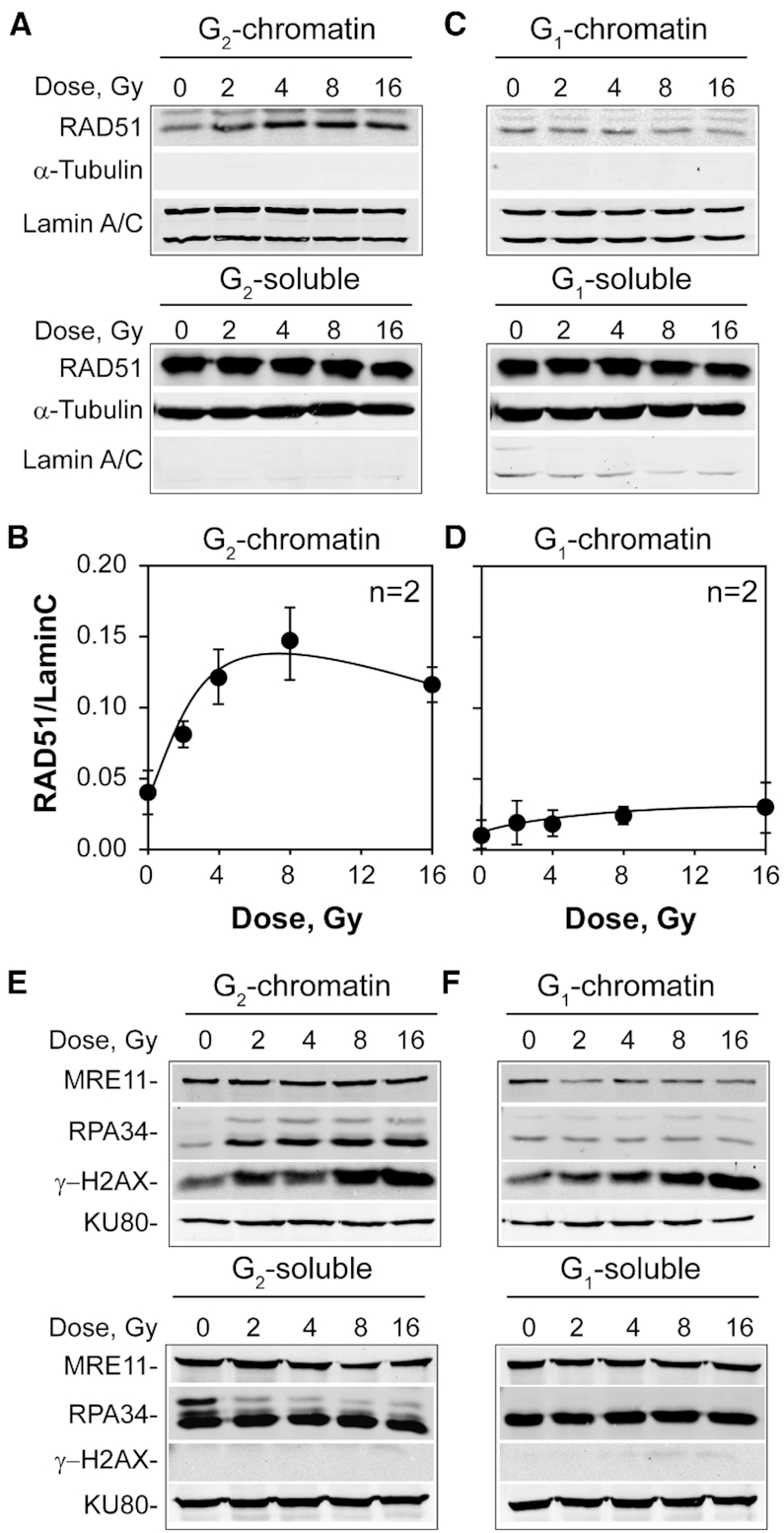
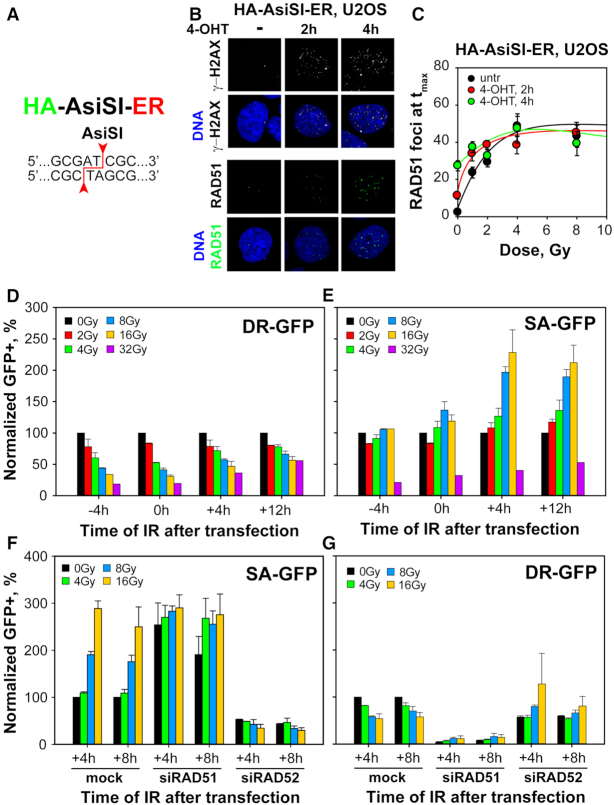
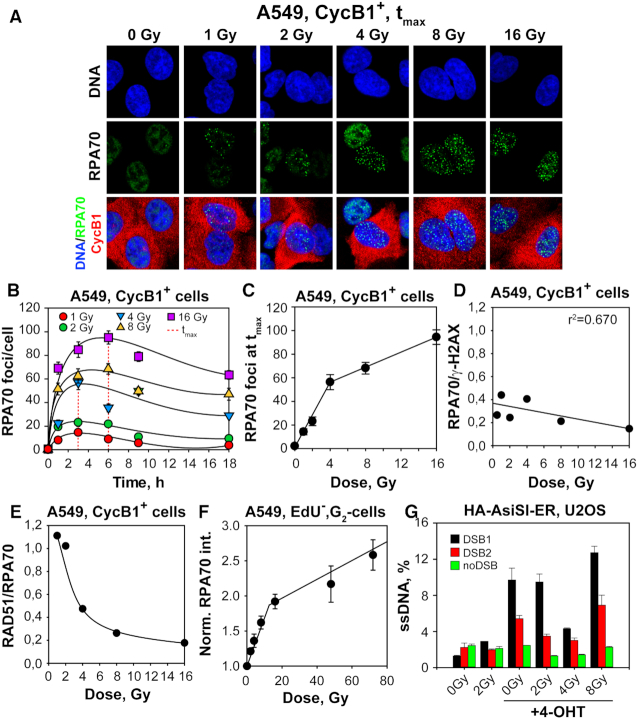
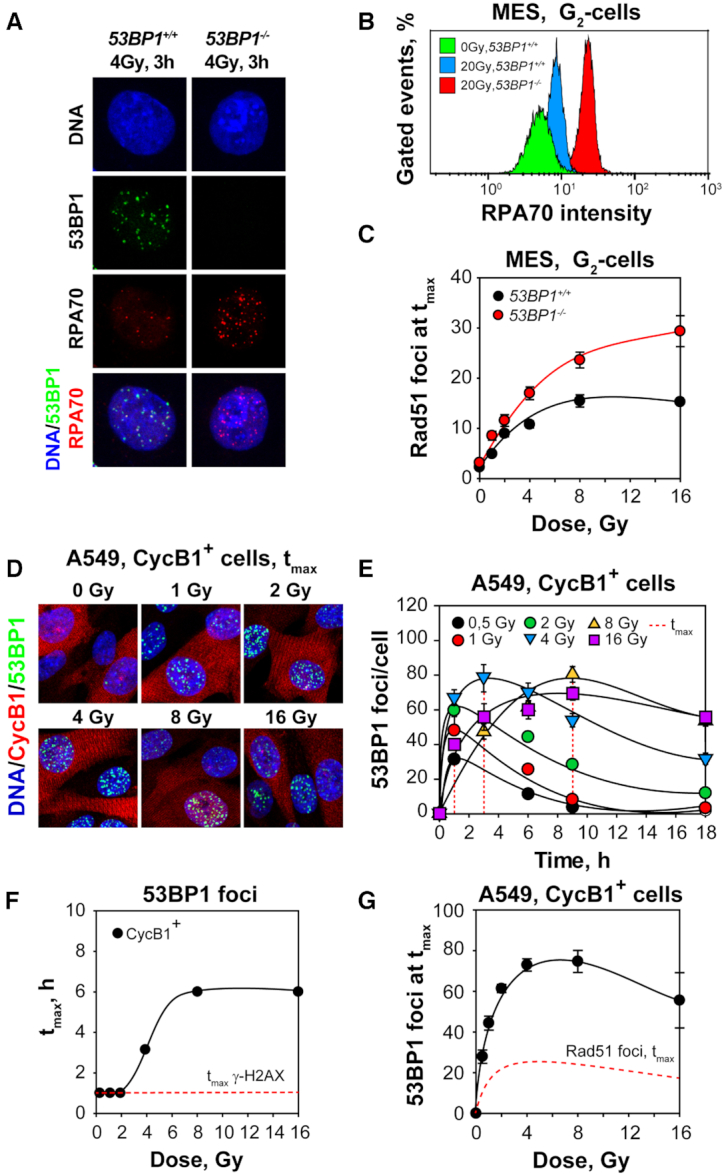
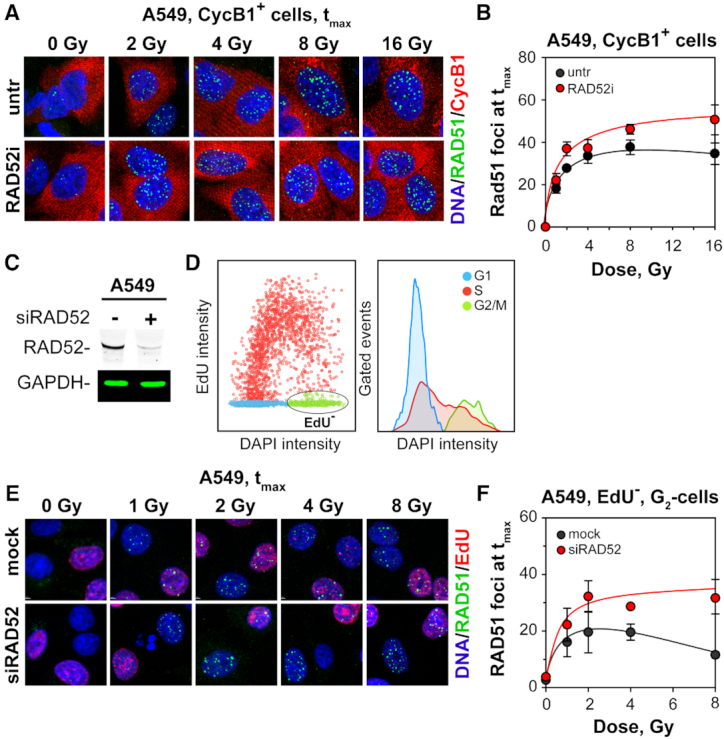
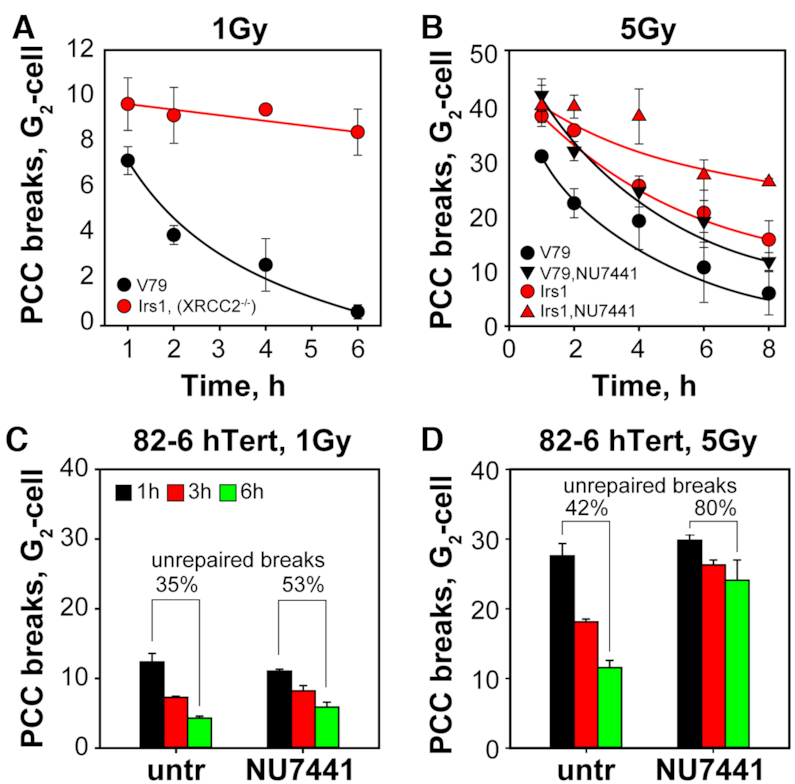
In vertebrates, genomic DNA double-strand breaks (DSBs) are removed by non-homologous end-joining processes: classical non-homologous end-joining (c-NHEJ) and alternative end-joining (alt-EJ); or by homology-dependent processes: gene-conversion (GC) and single-strand annealing (SSA). Surprisingly, these repair pathways are not real alternative options restoring genome integrity with equal efficiency, but show instead striking differences in speed, accuracy and cell-cycle-phase dependence. As a consequence, engagement of one pathway may be associated with processing-risks for the genome absent from another pathway. Characterization of engagement-parameters and their consequences is, therefore, essential for understanding effects on the genome of DSB-inducing agents, such as ionizing-radiation (IR). Here, by addressing pathway selection in G2-phase, we discover regulatory confinements in GC with consequences for SSA- and c-NHEJ-engagement. We show pronounced suppression of GC with increasing DSB-load that is not due to RAD51 availability and which is delimited but not defined by 53BP1 and RAD52. Strikingly, at low DSB-loads, GC repairs ∼50% of DSBs, whereas at high DSB-loads its contribution is undetectable. Notably, with increasing DSB-load and the associated suppression of GC, SSA gains ground, while alt-EJ is suppressed. These observations explain earlier, apparently contradictory results and advance our understanding of logic and mechanisms underpinning the wiring between DSB repair pathways.
© The Author(s) 2019. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Chromosome breaks generated by low doses of ionizing radiation in G2-phase are processed exclusively by gene conversion.DNA Repair (Amst). 2020 May;89:102828. doi: 10.1016/j.dnarep.2020.102828. Epub 2020 Feb 27. DNA Repair (Amst). 2020. PMID: 32143127
-
Analysis of chromatid-break-repair detects a homologous recombination to non-homologous end-joining switch with increasing load of DNA double-strand breaks.Mutat Res Genet Toxicol Environ Mutagen. 2021 Jul;867:503372. doi: 10.1016/j.mrgentox.2021.503372. Epub 2021 Jun 12. Mutat Res Genet Toxicol Environ Mutagen. 2021. PMID: 34266628
-
Regulation of DNA repair in the absence of classical non-homologous end joining.DNA Repair (Amst). 2018 Aug;68:34-40. doi: 10.1016/j.dnarep.2018.06.001. Epub 2018 Jun 12. DNA Repair (Amst). 2018. PMID: 29929045
-
New Facets of DNA Double Strand Break Repair: Radiation Dose as Key Determinant of HR versus c-NHEJ Engagement.Int J Mol Sci. 2023 Oct 6;24(19):14956. doi: 10.3390/ijms241914956. Int J Mol Sci. 2023. PMID: 37834403 Free PMC article. Review.
-
Mechanisms of DNA double strand break repair and chromosome aberration formation.Cytogenet Genome Res. 2004;104(1-4):14-20. doi: 10.1159/000077461. Cytogenet Genome Res. 2004. PMID: 15162010 Review.
Cited by
-
Increased Gene Targeting in Hyper-Recombinogenic LymphoBlastoid Cell Lines Leaves Unchanged DSB Processing by Homologous Recombination.Int J Mol Sci. 2022 Aug 16;23(16):9180. doi: 10.3390/ijms23169180. Int J Mol Sci. 2022. PMID: 36012445 Free PMC article.
-
Tracking telomere fusions through crisis reveals conflict between DNA transcription and the DNA damage response.NAR Cancer. 2021 Mar;3(1):zcaa044. doi: 10.1093/narcan/zcaa044. Epub 2021 Jan 6. NAR Cancer. 2021. PMID: 33447828 Free PMC article.
-
Shift in G1-Checkpoint from ATM-Alone to a Cooperative ATM Plus ATR Regulation with Increasing Dose of Radiation.Cells. 2021 Dec 27;11(1):63. doi: 10.3390/cells11010063. Cells. 2021. PMID: 35011623 Free PMC article.
-
The p38/MK2 Pathway Functions as Chk1-Backup Downstream of ATM/ATR in G2-Checkpoint Activation in Cells Exposed to Ionizing Radiation.Cells. 2023 May 14;12(10):1387. doi: 10.3390/cells12101387. Cells. 2023. PMID: 37408221 Free PMC article.
-
PTEN Loss Enhances Error-Prone DSB Processing and Tumor Cell Radiosensitivity by Suppressing RAD51 Expression and Homologous Recombination.Int J Mol Sci. 2022 Oct 25;23(21):12876. doi: 10.3390/ijms232112876. Int J Mol Sci. 2022. PMID: 36361678 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
