

Fragment-based screening identifies molecules targeting the substrate-binding ankyrin repeat domains of tankyrase
- PMID: 31836723
- PMCID: PMC6911004
- DOI: 10.1038/s41598-019-55240-5
Fragment-based screening identifies molecules targeting the substrate-binding ankyrin repeat domains of tankyrase
Abstract
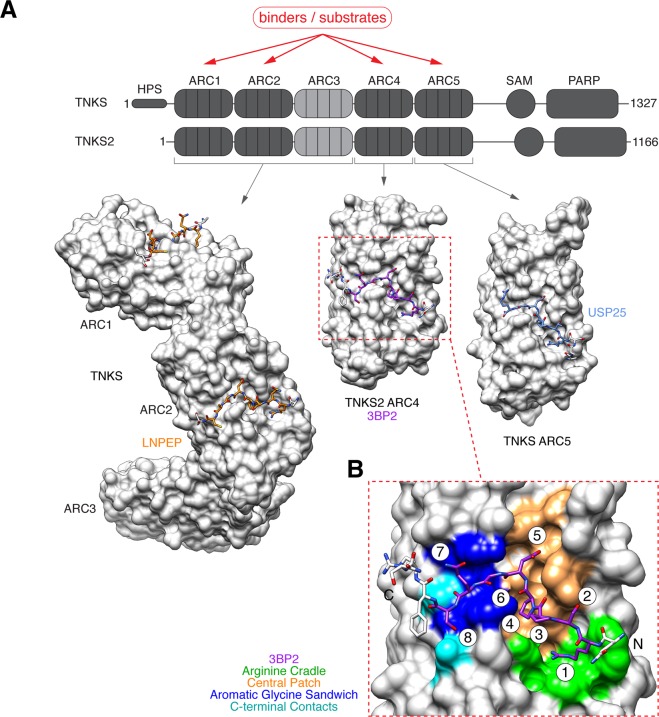
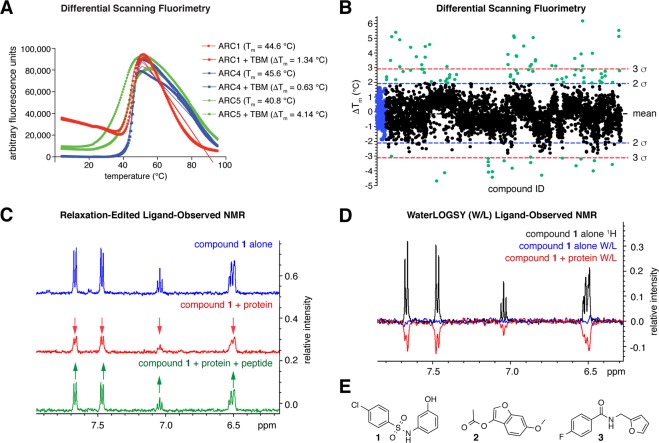
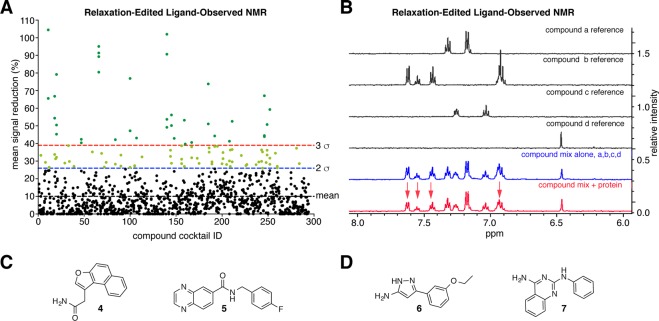
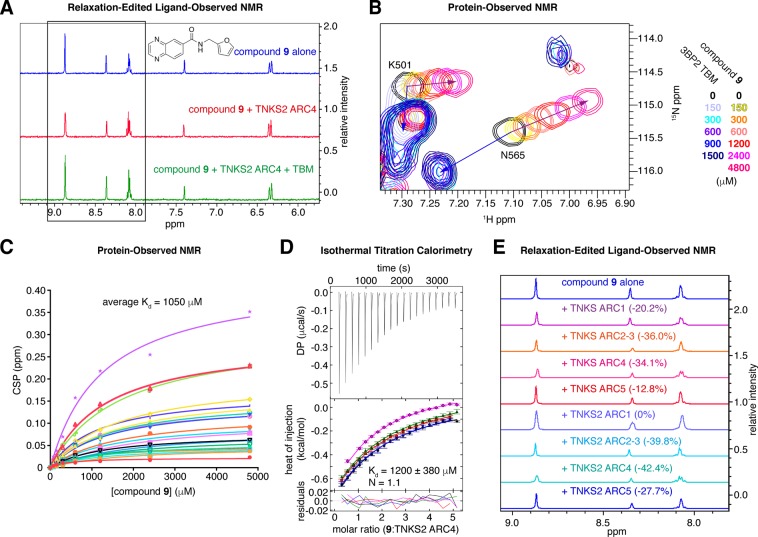
The PARP enzyme and scaffolding protein tankyrase (TNKS, TNKS2) uses its ankyrin repeat clusters (ARCs) to bind a wide range of proteins and thereby controls diverse cellular functions. A number of these are implicated in cancer-relevant processes, including Wnt/β-catenin signalling, Hippo signalling and telomere maintenance. The ARCs recognise a conserved tankyrase-binding peptide motif (TBM). All currently available tankyrase inhibitors target the catalytic domain and inhibit tankyrase's poly(ADP-ribosyl)ation function. However, there is emerging evidence that catalysis-independent "scaffolding" mechanisms contribute to tankyrase function. Here we report a fragment-based screening programme against tankyrase ARC domains, using a combination of biophysical assays, including differential scanning fluorimetry (DSF) and nuclear magnetic resonance (NMR) spectroscopy. We identify fragment molecules that will serve as starting points for the development of tankyrase substrate binding antagonists. Such compounds will enable probing the scaffolding functions of tankyrase, and may, in the future, provide potential alternative therapeutic approaches to inhibiting tankyrase activity in cancer and other conditions.
Conflict of interest statement
The authors declare no competing interests.
Figures
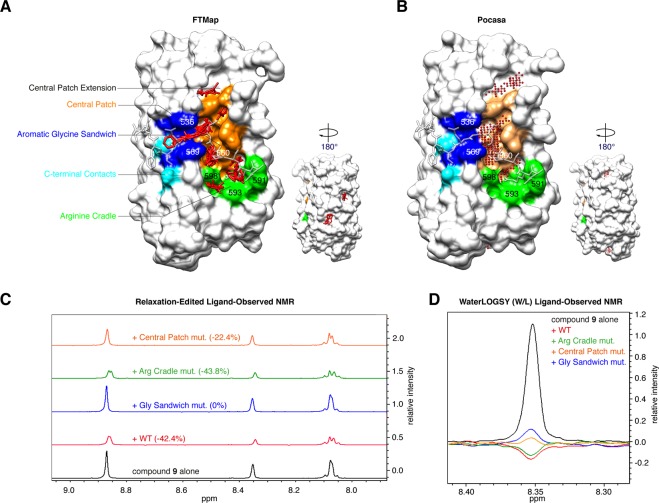
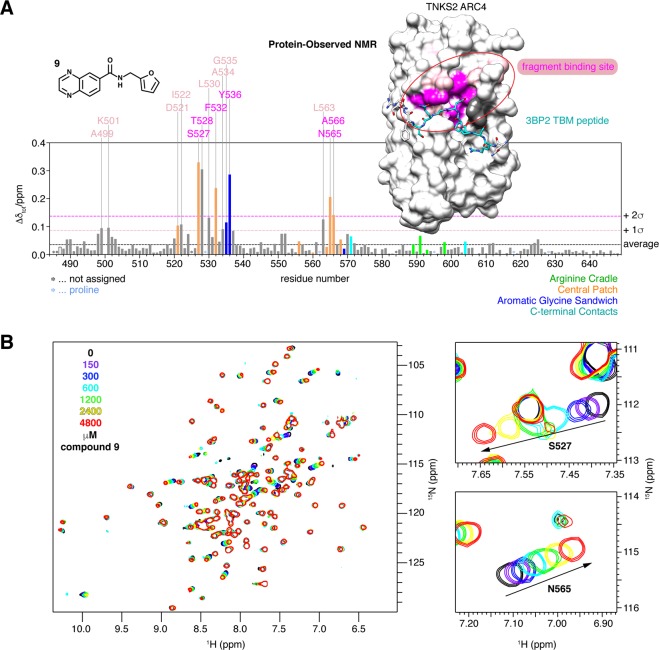
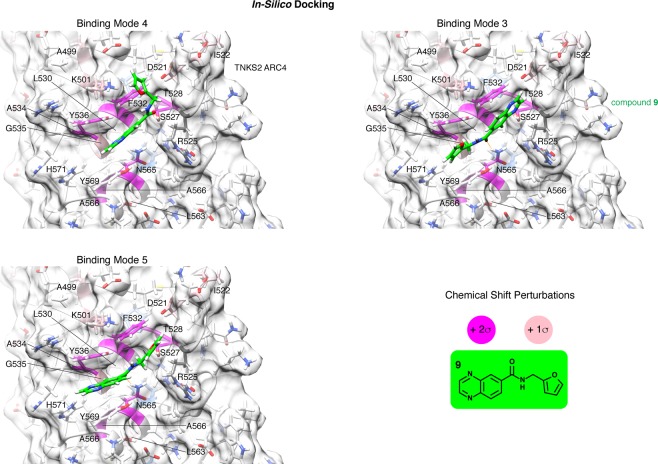
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
