

Retroviruses drive the rapid evolution of mammalian APOBEC3 genes
- PMID: 31843890
- PMCID: PMC6955324
- DOI: 10.1073/pnas.1914183116
Retroviruses drive the rapid evolution of mammalian APOBEC3 genes
Abstract
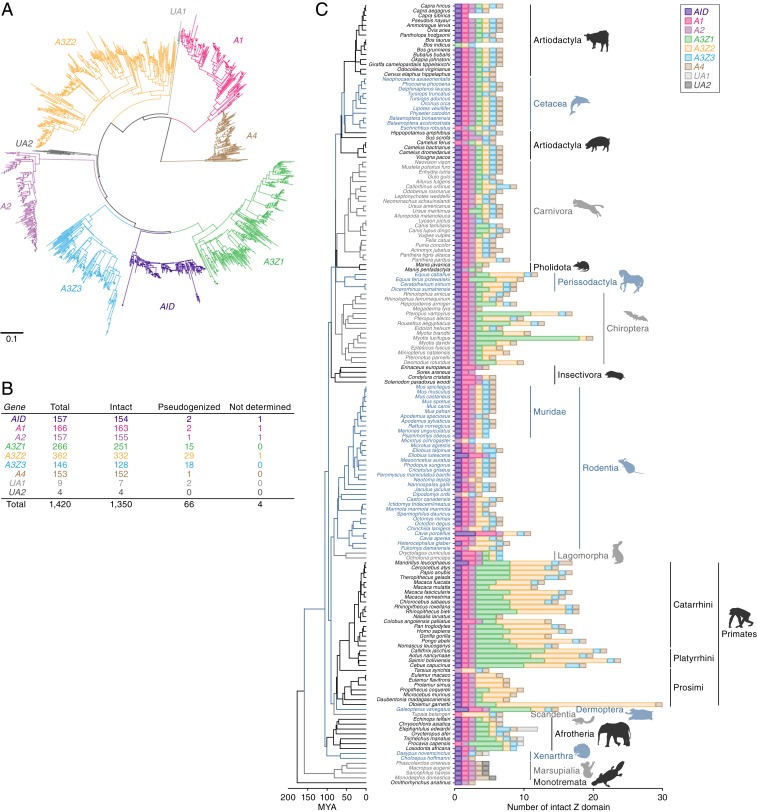
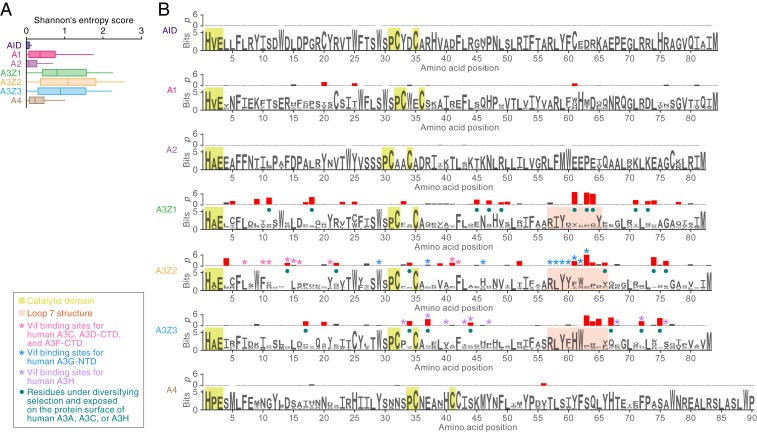
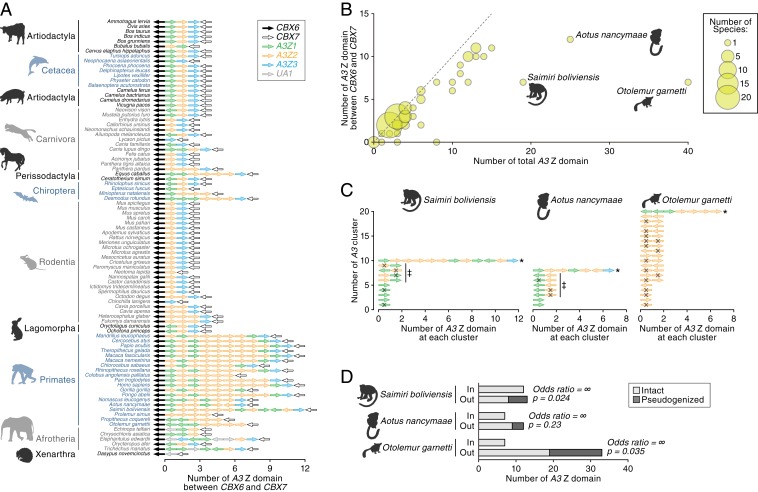
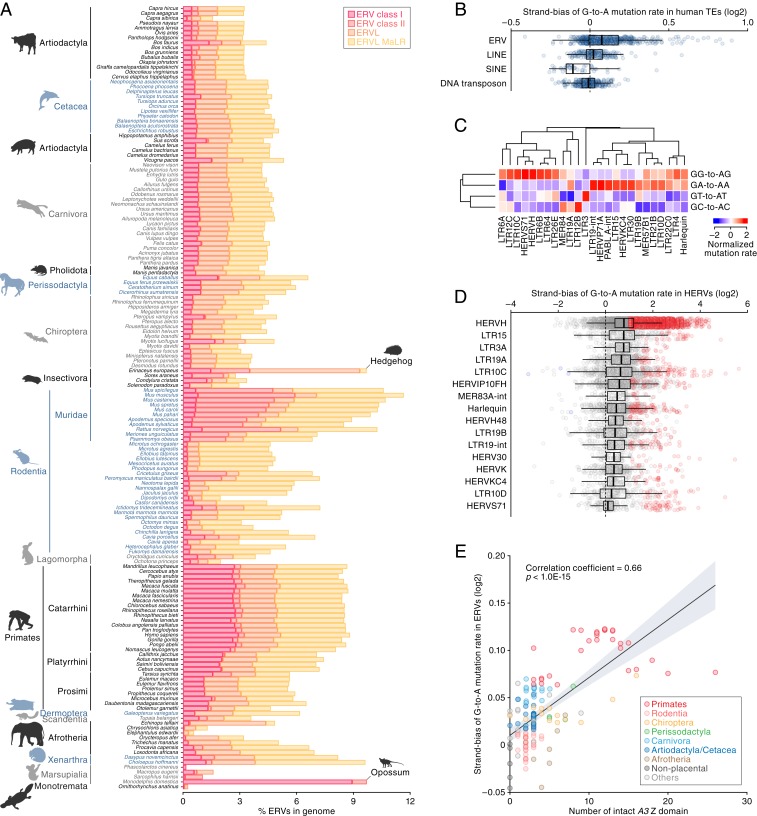
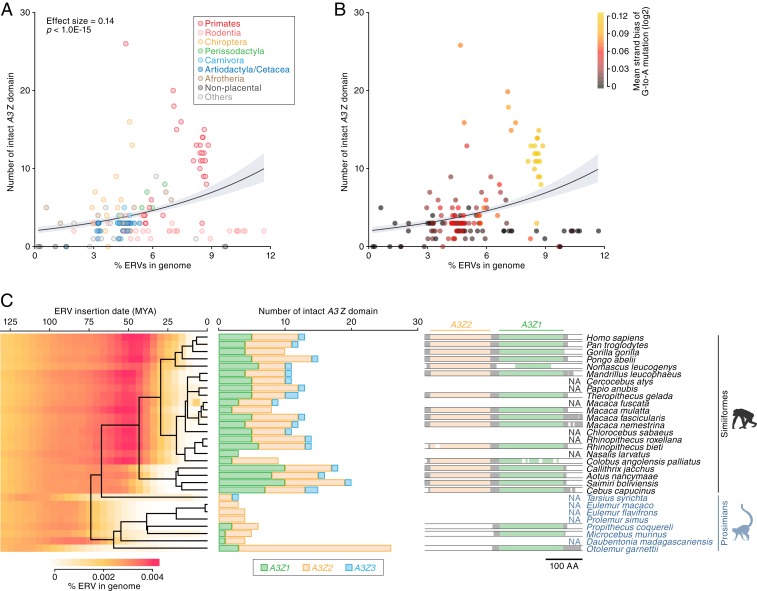
APOBEC3 (A3) genes are members of the AID/APOBEC gene family that are found exclusively in mammals. A3 genes encode antiviral proteins that restrict the replication of retroviruses by inducing G-to-A mutations in their genomes and have undergone extensive amplification and diversification during mammalian evolution. Endogenous retroviruses (ERVs) are sequences derived from ancient retroviruses that are widespread mammalian genomes. In this study we characterize the A3 repertoire and use the ERV fossil record to explore the long-term history of coevolutionary interaction between A3s and retroviruses. We examine the genomes of 160 mammalian species and identify 1,420 AID/APOBEC-related genes, including representatives of previously uncharacterized lineages. We show that A3 genes have been amplified in mammals and that amplification is positively correlated with the extent of germline colonization by ERVs. Moreover, we demonstrate that the signatures of A3-mediated mutation can be detected in ERVs found throughout mammalian genomes and show that in mammalian species with expanded A3 repertoires, ERVs are significantly enriched for G-to-A mutations. Finally, we show that A3 amplification occurred concurrently with prominent ERV invasions in primates. Our findings establish that conflict with retroviruses is a major driving force for the rapid evolution of mammalian A3 genes.
Keywords: APOBEC3; endogenous retrovirus; evolutionary arms race; gene amplification; mammal.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Conticello S. G., Langlois M. A., Yang Z., Neuberger M. S., DNA deamination in immunity: AID in the context of its APOBEC relatives. Adv. Immunol. 94, 37–73 (2007). - PubMed
-
- Teng B., Burant C. F., Davidson N. O., Molecular cloning of an apolipoprotein B messenger RNA editing protein. Science 260, 1816–1819 (1993). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
