

Characterization of splice-altering mutations in inherited predisposition to cancer
- PMID: 31843900
- PMCID: PMC6936554
- DOI: 10.1073/pnas.1915608116
Characterization of splice-altering mutations in inherited predisposition to cancer
Abstract
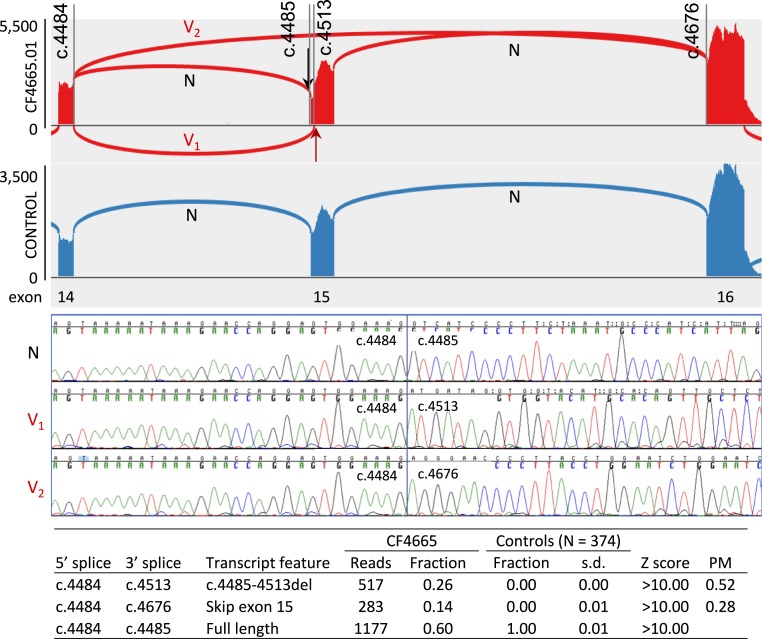
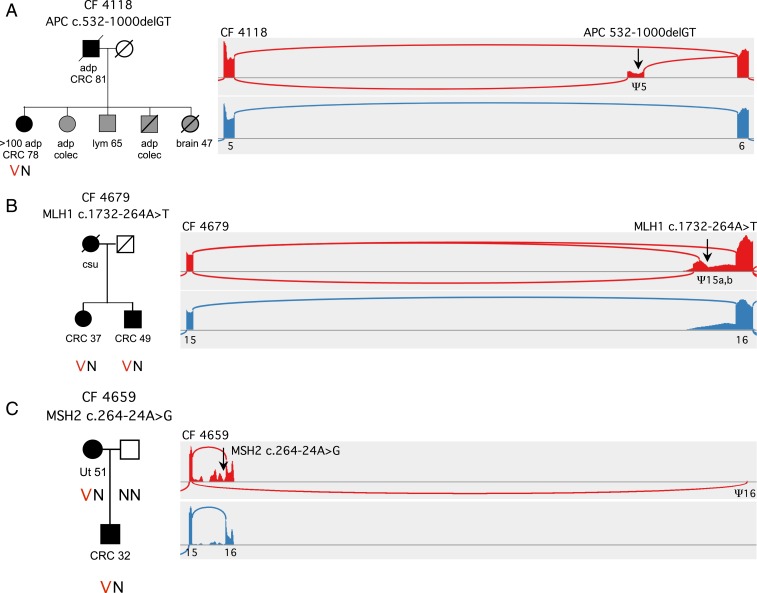
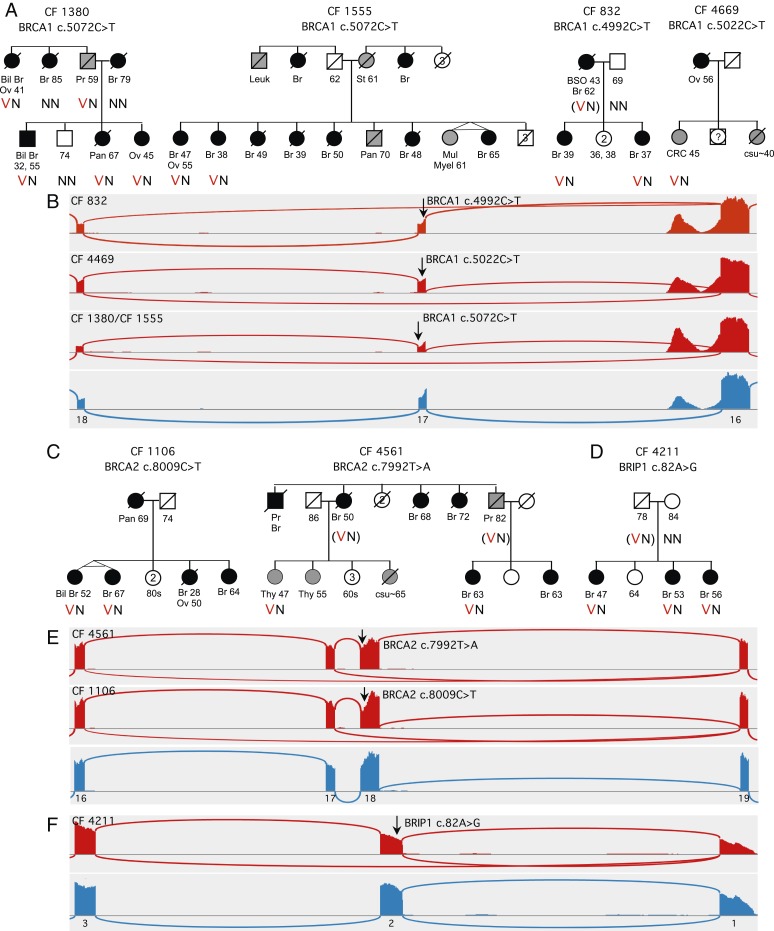
Mutations responsible for inherited disease may act by disrupting normal transcriptional splicing. Such mutations can be difficult to detect, and their effects difficult to characterize, because many lie deep within exons or introns where they may alter splice enhancers or silencers or introduce new splice acceptors or donors. Multiple mutation-specific and genome-wide approaches have been developed to evaluate these classes of mutations. We introduce a complementary experimental approach, cBROCA, which yields qualitative and quantitative assessments of the effects of genomic mutations on transcriptional splicing of tumor suppressor genes. cBROCA analysis is undertaken by deriving complementary DNA (cDNA) from puromycin-treated patient lymphoblasts, hybridizing the cDNA to the BROCA panel of tumor suppressor genes, and then multiplex sequencing to very high coverage. At each splice junction suggested by split sequencing reads, read depths of test and control samples are compared. Significant Z scores indicate altered transcripts, over and above naturally occurring minor transcripts, and comparisons of read depths indicate relative abundances of mutant and normal transcripts. BROCA analysis of genomic DNA suggested 120 rare mutations from 150 families with cancers of the breast, ovary, uterus, or colon, in >600 informative genotyped relatives. cBROCA analysis of their transcripts revealed a wide variety of consequences of abnormal splicing in tumor suppressor genes, including whole or partial exon skipping, exonification of intronic sequence, loss or gain of exonic and intronic splicing enhancers and silencers, complete intron retention, hypomorphic alleles, and combinations of these alterations. Combined with pedigree analysis, cBROCA sequencing contributes to understanding the clinical consequences of rare inherited mutations.
Keywords: cancer; mutation; splicing.
Conflict of interest statement
Competing interest statement: R.O. is an employee of Color Genomics. Z.T. is an employee of Color Genomics. T.W. consults for Color Genomics.
Figures


References
Grants and funding
LinkOut - more resources
Full Text Sources
