

Loss of ADAMTS19 causes progressive non-syndromic heart valve disease
- PMID: 31844321
- PMCID: PMC7197892
- DOI: 10.1038/s41588-019-0536-2
Loss of ADAMTS19 causes progressive non-syndromic heart valve disease
Abstract
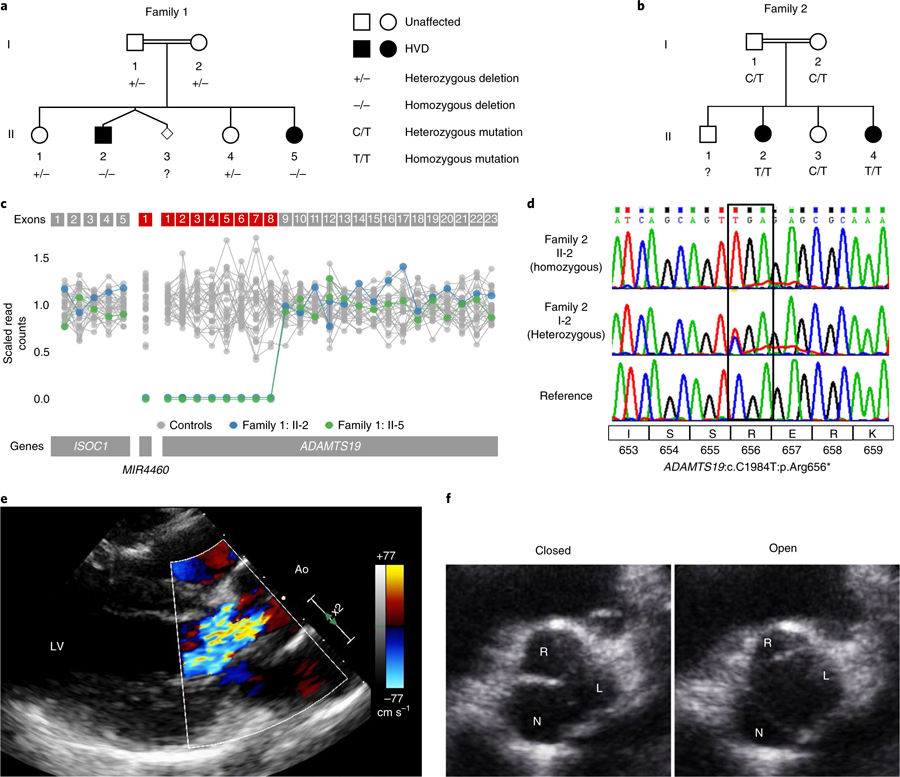
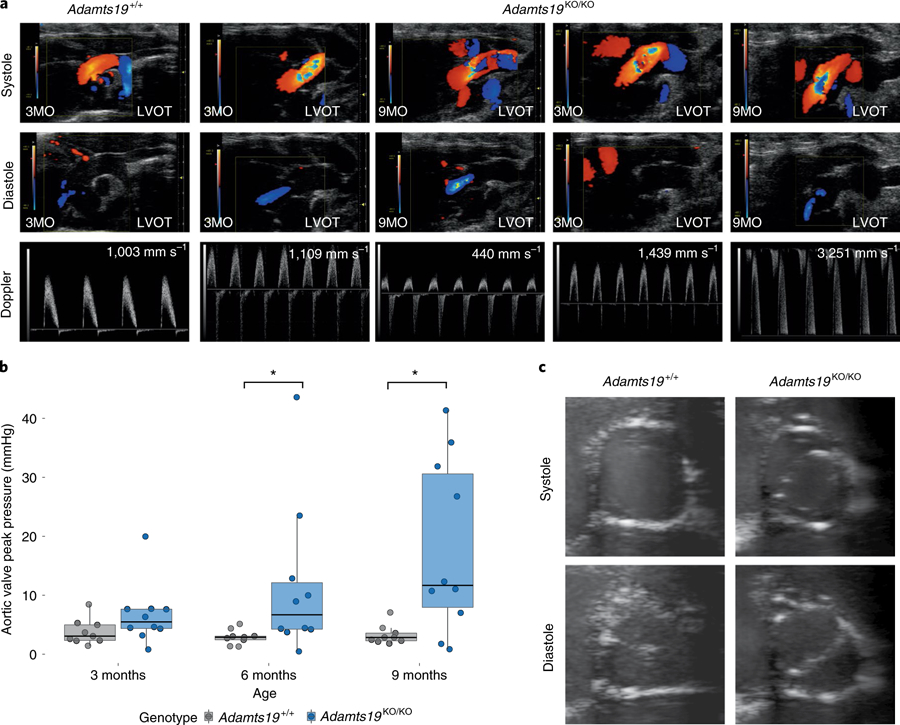
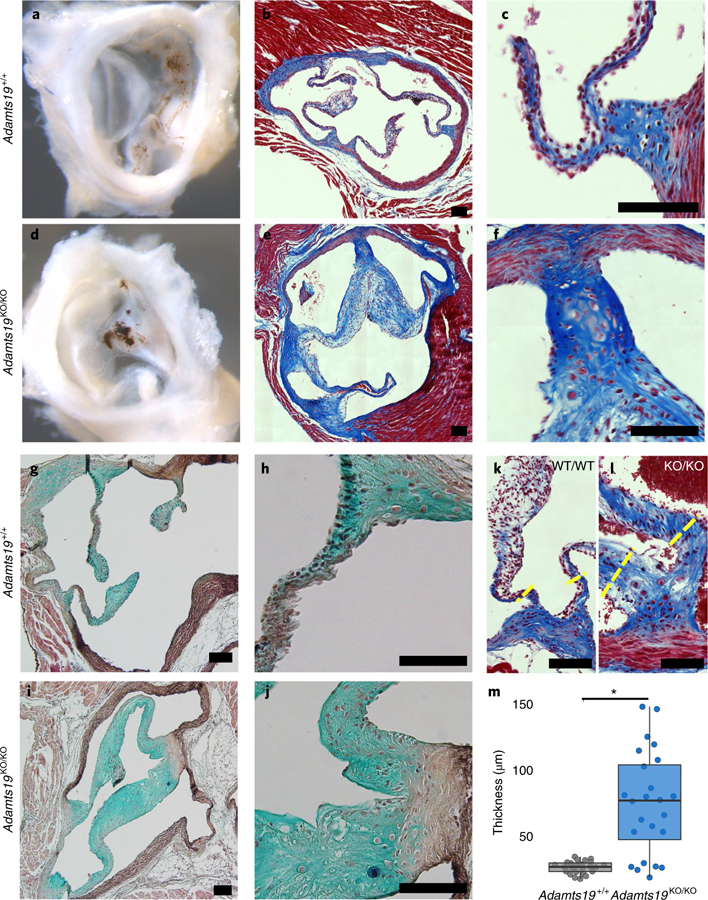
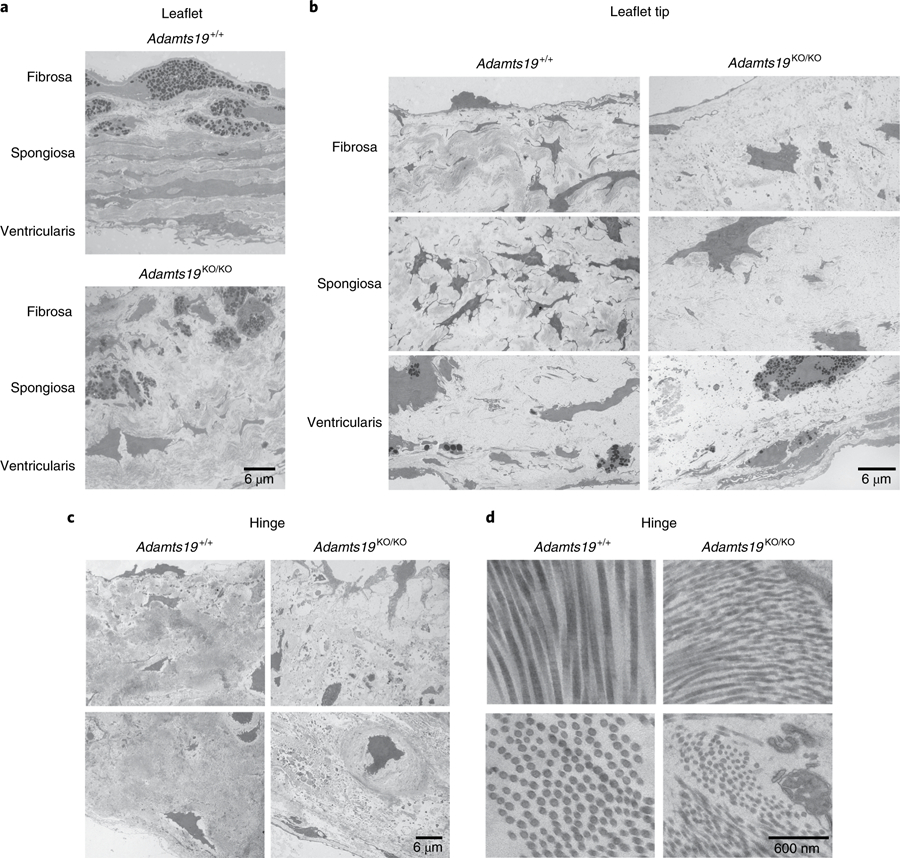
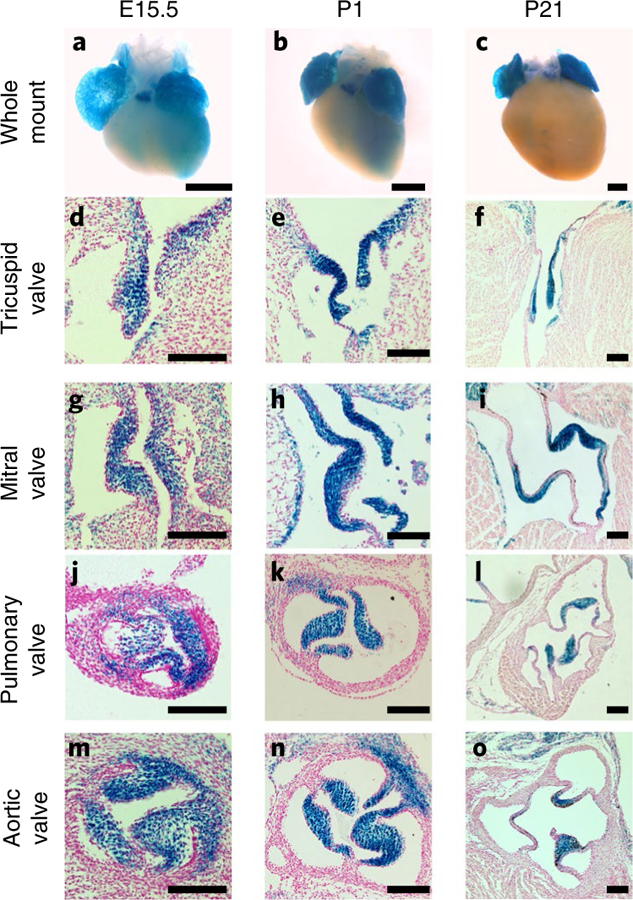
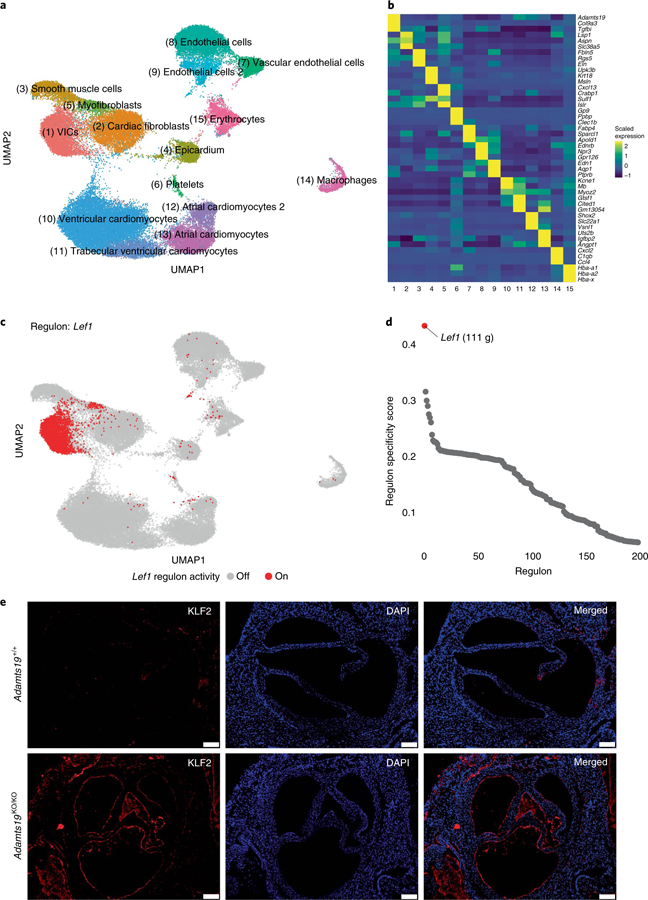
Valvular heart disease is observed in approximately 2% of the general population1. Although the initial observation is often localized (for example, to the aortic or mitral valve), disease manifestations are regularly observed in the other valves and patients frequently require surgery. Despite the high frequency of heart valve disease, only a handful of genes have so far been identified as the monogenic causes of disease2-7. Here we identify two consanguineous families, each with two affected family members presenting with progressive heart valve disease early in life. Whole-exome sequencing revealed homozygous, truncating nonsense alleles in ADAMTS19 in all four affected individuals. Homozygous knockout mice for Adamts19 show aortic valve dysfunction, recapitulating aspects of the human phenotype. Expression analysis using a lacZ reporter and single-cell RNA sequencing highlight Adamts19 as a novel marker for valvular interstitial cells; inference of gene regulatory networks in valvular interstitial cells positions Adamts19 in a highly discriminatory network driven by the transcription factor lymphoid enhancer-binding factor 1 downstream of the Wnt signaling pathway. Upregulation of endocardial Krüppel-like factor 2 in Adamts19 knockout mice precedes hemodynamic perturbation, showing that a tight balance in the Wnt-Adamts19-Klf2 axis is required for proper valve maturation and maintenance.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
-
- Nkomo VT et al. Burden of valvular heart diseases: a population-based study. Lancet 368, 1005–1011 (2006). - PubMed
-
- Garg V et al. Mutations in NOTCH1 cause aortic valve disease. Nature 437, 270–274 (2005). - PubMed
-
- Padang R, Bagnall RD, Richmond DR, Bannon PG & Semsarian C Rare non-synonymous variations in the transcriptional activation domains of GATA5 in bicuspid aortic valve disease. J. Mol. Cell. Cardiol 53, 277–281 (2012). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
