

Elucidating the Role of K+ Channels during In Vitro Capacitation of Boar Spermatozoa: Do SLO1 Channels Play a Crucial Role?
- PMID: 31847486
- PMCID: PMC6940911
- DOI: 10.3390/ijms20246330
Elucidating the Role of K+ Channels during In Vitro Capacitation of Boar Spermatozoa: Do SLO1 Channels Play a Crucial Role?
Abstract
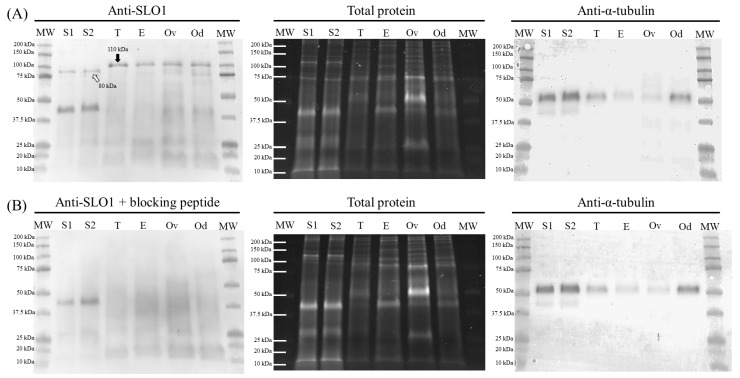
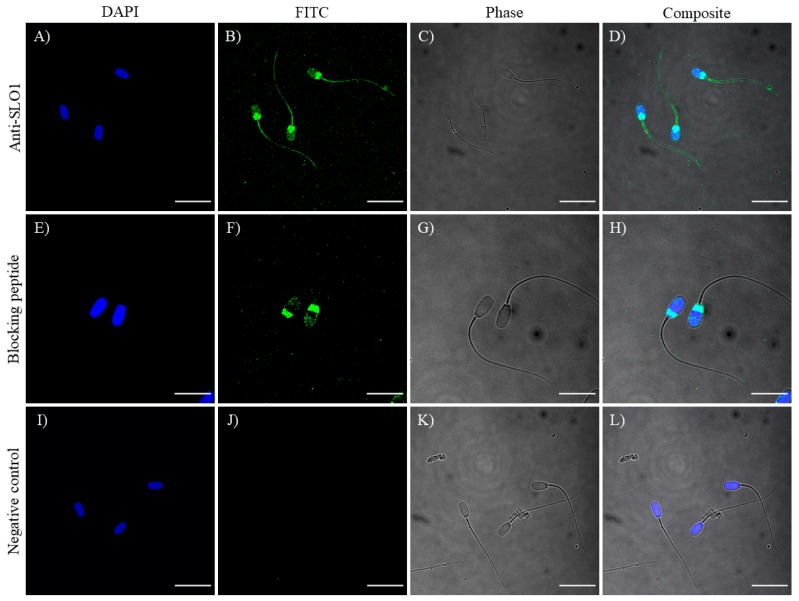
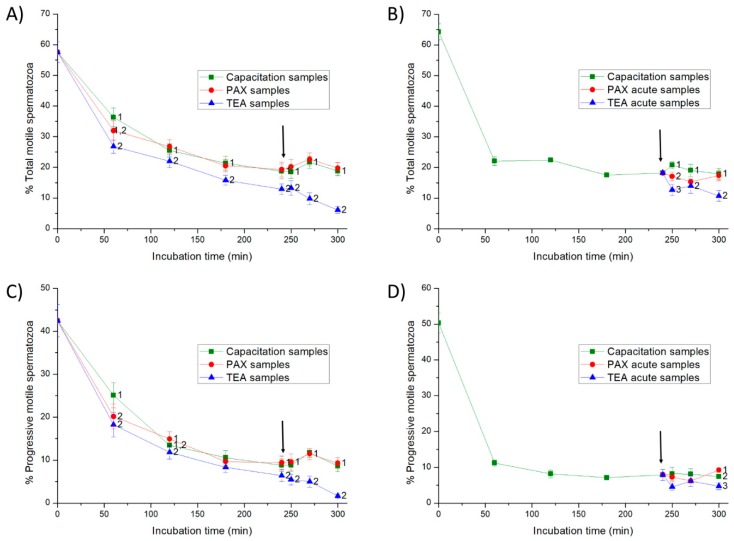
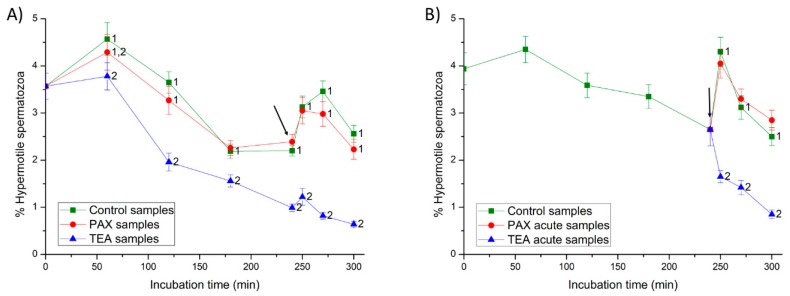
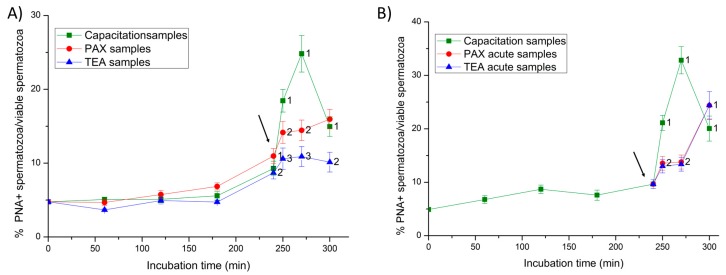
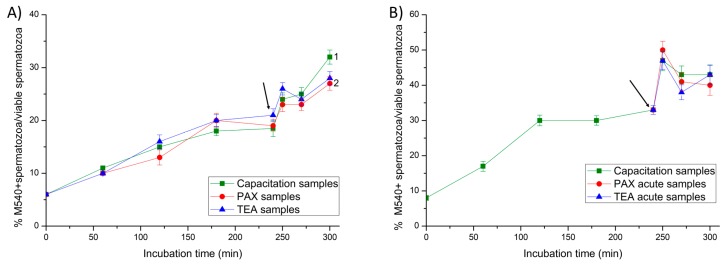
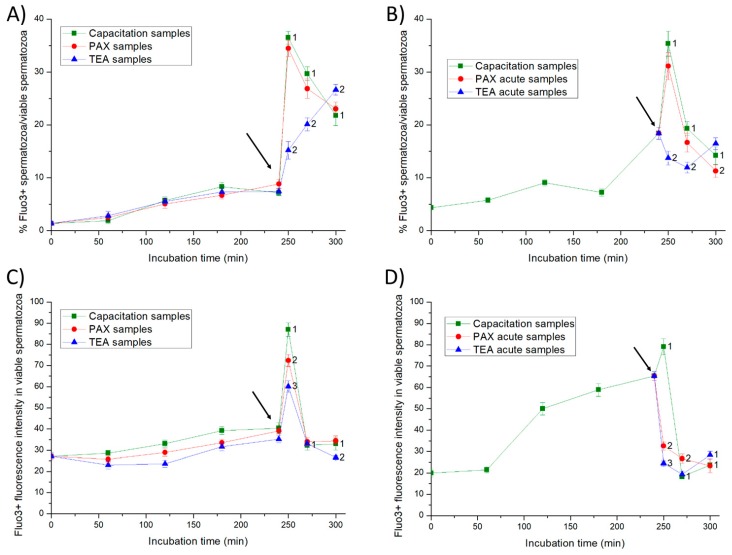
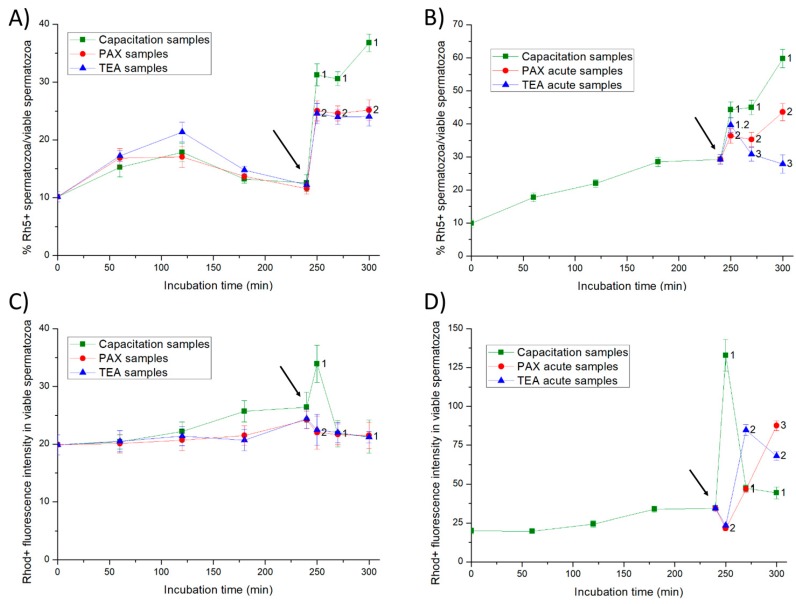
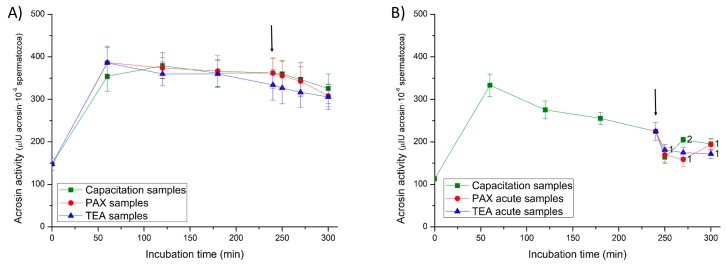
This study sought to identify and localize SLO1 channels in boar spermatozoa by immunoblotting and immunofluorescence, and to determine their physiological role during in vitro sperm capacitation. Sperm samples from 14 boars were incubated in a capacitation medium for 300 min in the presence of paxilline (PAX), a specific SLO1-channel blocker, added either at 0 min or after 240 min of incubation. Negative controls were incubated in capacitation medium, and positive controls in capacitation medium plus tetraethyl ammonium (TEA), a general K+-channel blocker, also added at 0 min or after 240 min of incubation. In all samples, acrosome exocytosis was triggered with progesterone after 240 min of incubation. Sperm motility and kinematics, integrity of plasma and acrosome membranes, membrane lipid disorder, intracellular calcium levels and acrosin activity were evaluated after 0, 60, 120, 180, 240, 250, 270 and 300 min of incubation. In boar spermatozoa, SLO1 channels were found to have 80 kDa and be localized in the anterior postacrosomal region and the mid and principal piece of the tail; their specific blockage through PAX resulted in altered calcium levels and acrosome exocytosis. As expected, TEA blocker impaired in vitro sperm capacitation, by altering sperm motility and kinematics and calcium levels. In conclusion, SLO1 channels are crucial for the acrosome exocytosis induced by progesterone in in vitro capacitated boar spermatozoa.
Keywords: PAX; SLO1 channels; TEA; boar spermatozoa; in vitro capacitation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- De la Vega-Beltran J.L., Sánchez-Cárdenas C., Krapf D., Hernandez-González E.O., Wertheimer E., Treviño C.L., Visconti P.E., Darszon A. Mouse sperm membrane potential hyperpolarization is necessary and sufficient to prepare sperm for the acrosome reaction. J. Biol. Chem. 2012;287:44384–44393. doi: 10.1074/jbc.M112.393488. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
