

Evolution of a multifunctional trait: shared effects of foraging ecology and thermoregulation on beak morphology, with consequences for song evolution
- PMID: 31847778
- PMCID: PMC6939928
- DOI: 10.1098/rspb.2019.2474
Evolution of a multifunctional trait: shared effects of foraging ecology and thermoregulation on beak morphology, with consequences for song evolution
Abstract
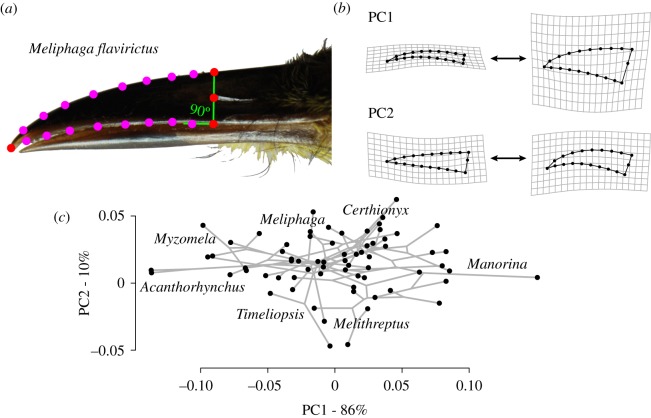
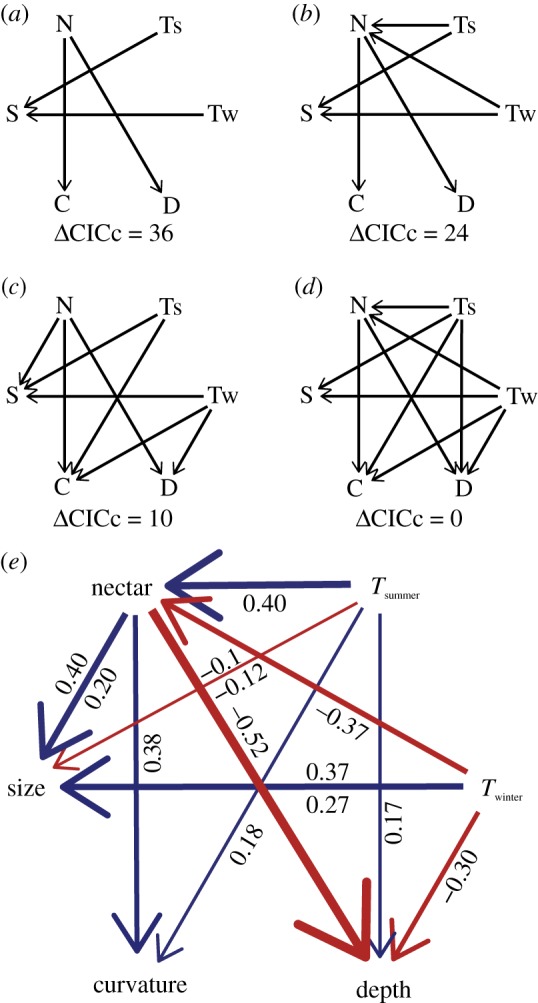
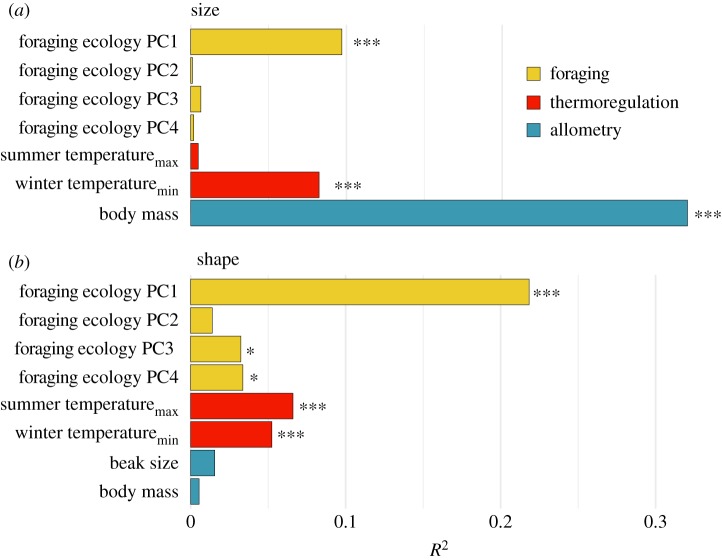
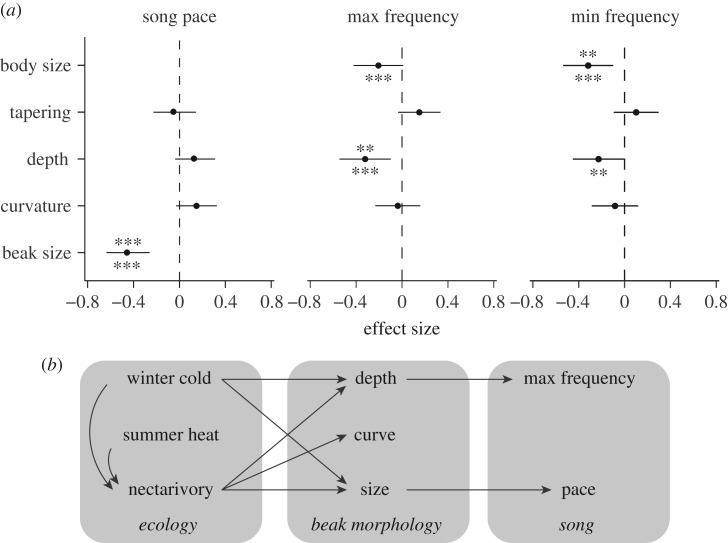
While morphological traits are often associated with multiple functions, it remains unclear how evolution balances the selective effects of different functions. Birds' beaks function not only in foraging but also in thermoregulating and singing, among other behaviours. Studies of beak evolution abound, however, most focus on a single function. Hence, we quantified relative contributions of different functions over an evolutionary timescale. We measured beak shape using geometric morphometrics and compared this trait with foraging behaviour, climatic variables and song characteristics in a phylogenetic comparative study of an Australasian radiation of songbirds (Meliphagidae). We found that both climate and foraging behaviour were significantly correlated with the beak shape and size. However, foraging ecology had a greater effect on shape, and climate had a nearly equal effect on size. We also found that evolutionary changes in beak morphology had significant consequences for vocal performance: species with elongate-shaped beaks sang at higher frequencies, while species with large beaks sang at a slower pace. The evolution of the avian beak exemplifies how morphological traits can be an evolutionary compromise among functions, and suggests that specialization along any functional axis may increase ecological divergence or reproductive isolation along others.
Keywords: Meliphagidae; beak shape; bird song; foraging; thermoregulation; trade-off.
Conflict of interest statement
We declare we have no competing interests
Figures
References
-
- Arnold SJ. 1983. Morphology, performance, and fitness. Am. Zool. 23, 347–361. (10.1093/icb/23.2.347) - DOI
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
