

The conserved C-terminus of Sss1p is required to maintain the endoplasmic reticulum permeability barrier
- PMID: 31848225
- PMCID: PMC7029116
- DOI: 10.1074/jbc.RA119.010378
The conserved C-terminus of Sss1p is required to maintain the endoplasmic reticulum permeability barrier
Abstract
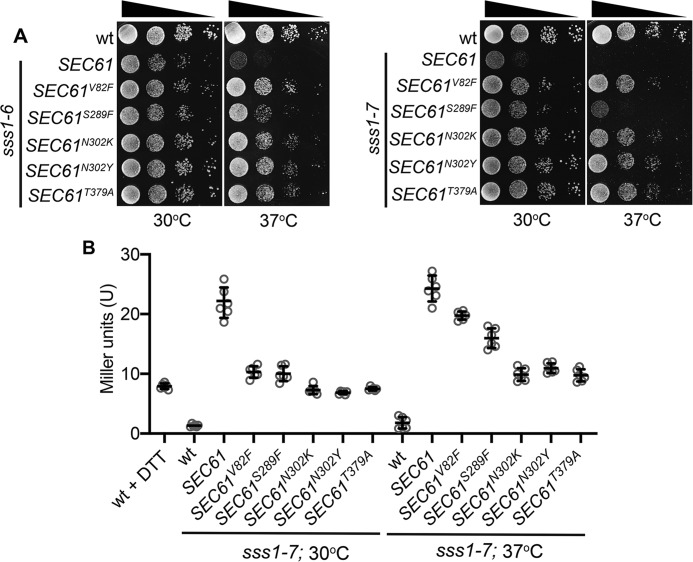
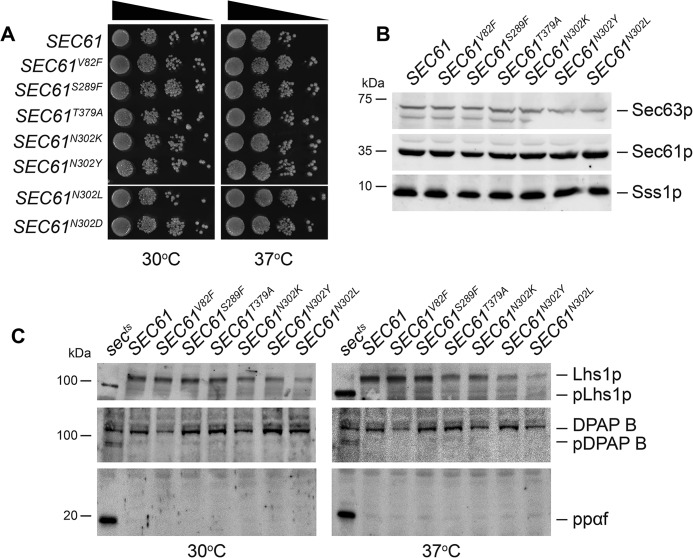
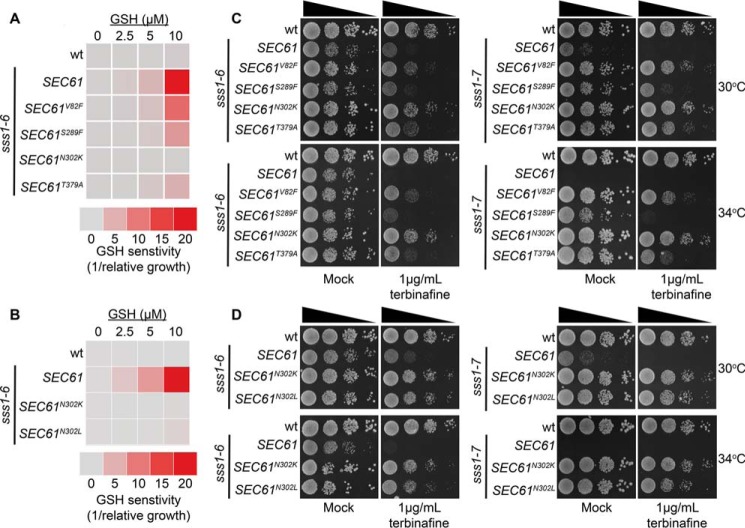
The endoplasmic reticulum (ER) is the entry point to the secretory pathway and major site of protein biogenesis. Translocation of secretory and integral membrane proteins across or into the ER membrane occurs via the evolutionarily conserved Sec61 complex, a heterotrimeric channel that comprises the Sec61p/Sec61α, Sss1p/Sec61γ, and Sbh1p/Sec61β subunits. In addition to forming a protein-conducting channel, the Sec61 complex also functions to maintain the ER permeability barrier, preventing the mass free flow of essential ER-enriched molecules and ions. Loss in Sec61 integrity is detrimental and implicated in the progression of disease. The Sss1p/Sec61γ C terminus is juxtaposed to the key gating module of Sec61p/Sec61α, and we hypothesize it is important for gating the ER translocon. The ER stress response was found to be constitutively induced in two temperature-sensitive sss1 mutants (sss1ts ) that are still proficient to conduct ER translocation. A screen to identify intergenic mutations that allow for sss1ts cells to grow at 37 °C suggests the ER permeability barrier to be compromised in these mutants. We propose the extreme C terminus of Sss1p/Sec61γ is an essential component of the gating module of the ER translocase and is required to maintain the ER permeability barrier.
Keywords: Sec61 complex; Sec61p/Sec61α; Sss1p/Sec61γ; endoplasmic reticulum (ER); endoplasmic reticulum stress (ER stress); endoplasmic reticulum–associated protein degradation (ERAD); gating; translocation; translocon gating.
© 2020 Witham et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article.
Figures





References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
