

Mass Spectrometry Based Immunopeptidomics Leads to Robust Predictions of Phosphorylated HLA Class I Ligands
- PMID: 31848261
- PMCID: PMC7000122
- DOI: 10.1074/mcp.TIR119.001641
Mass Spectrometry Based Immunopeptidomics Leads to Robust Predictions of Phosphorylated HLA Class I Ligands
Abstract
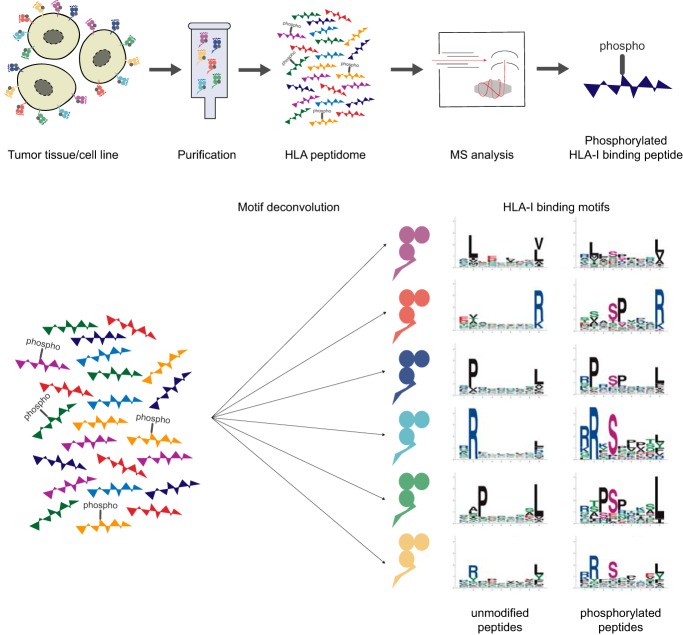
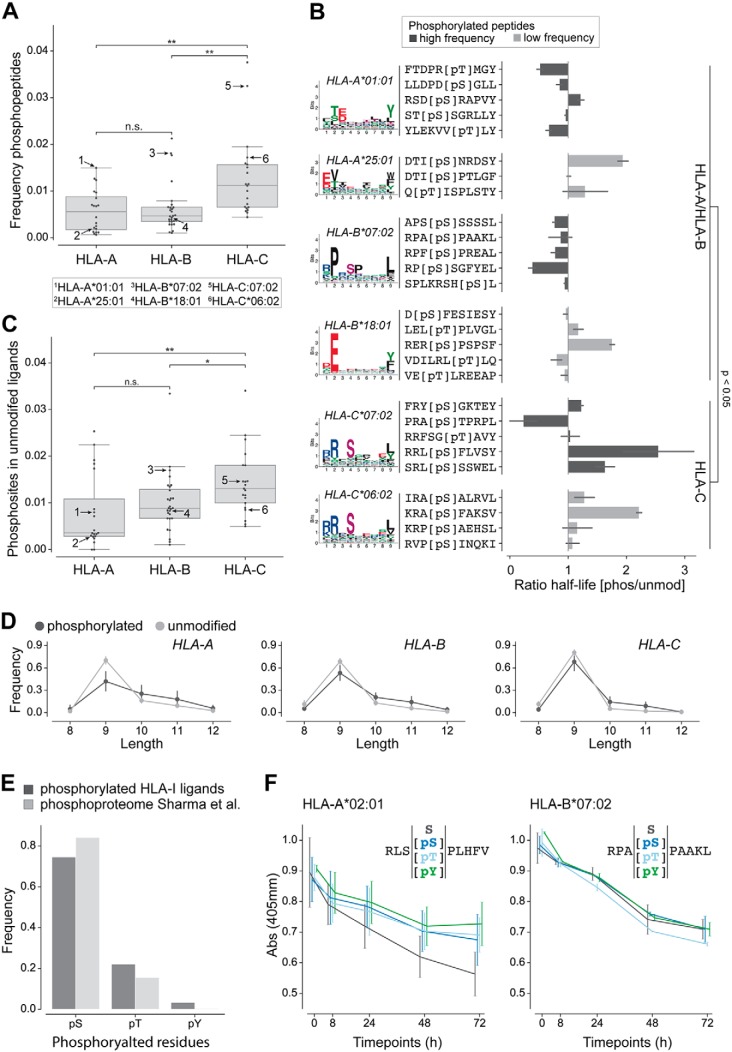
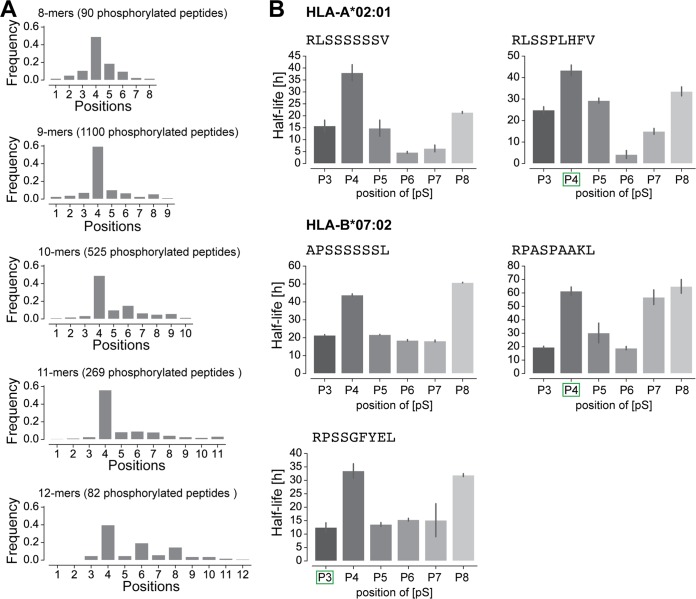
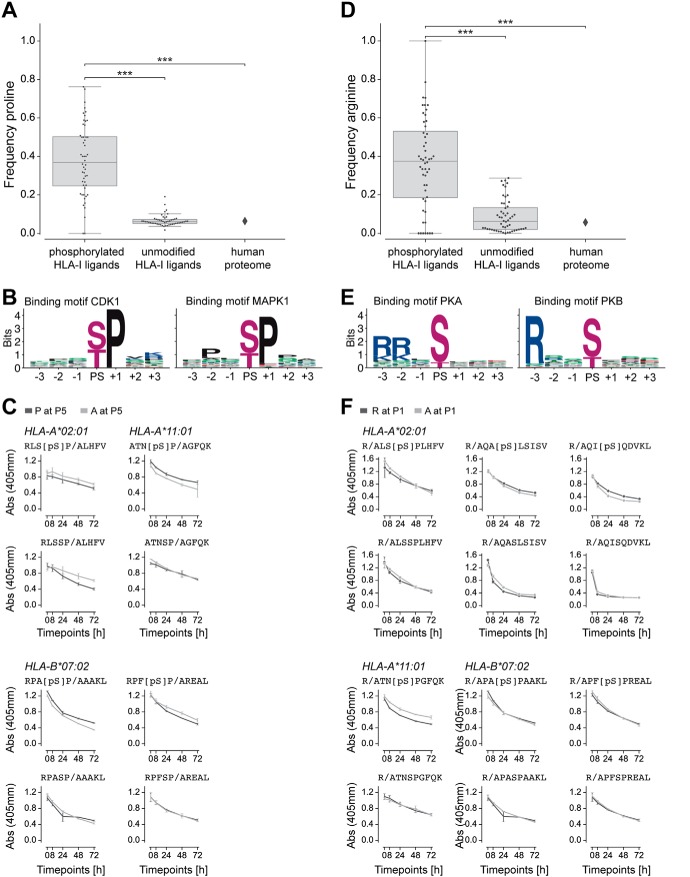
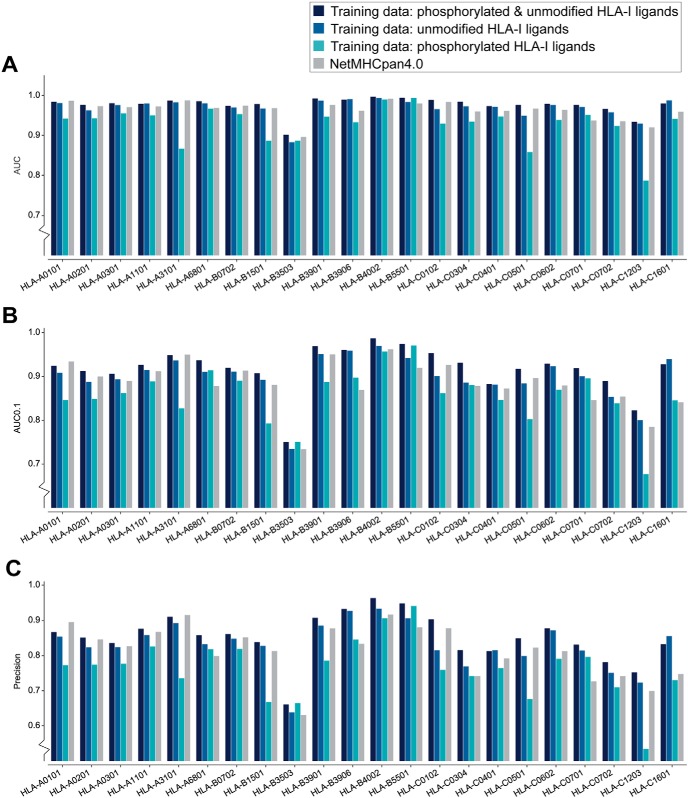
The presentation of peptides on class I human leukocyte antigen (HLA-I) molecules plays a central role in immune recognition of infected or malignant cells. In cancer, non-self HLA-I ligands can arise from many different alterations, including non-synonymous mutations, gene fusion, cancer-specific alternative mRNA splicing or aberrant post-translational modifications. Identifying HLA-I ligands remains a challenging task that requires either heavy experimental work for in vivo identification or optimized bioinformatics tools for accurate predictions. To date, no HLA-I ligand predictor includes post-translational modifications. To fill this gap, we curated phosphorylated HLA-I ligands from several immunopeptidomics studies (including six newly measured samples) covering 72 HLA-I alleles and retrieved a total of 2,066 unique phosphorylated peptides. We then expanded our motif deconvolution tool to identify precise binding motifs of phosphorylated HLA-I ligands. Our results reveal a clear enrichment of phosphorylated peptides among HLA-C ligands and demonstrate a prevalent role of both HLA-I motifs and kinase motifs on the presentation of phosphorylated peptides. These data further enabled us to develop and validate the first predictor of interactions between HLA-I molecules and phosphorylated peptides.
Keywords: HLA peptidomics; HLA-I ligand predictions; Mass spectrometry; computational biology; computational immunology; immunology; peptidomics; phosphorylated HLA-I binding motifs; phosphorylated HLA-I ligands; phosphorylation.
© 2020 Solleder et al.
Conflict of interest statement
* The authors declare that they have no conflicts of interest with the contents of this article.
Figures

References
-
- Neefjes J., Jongsma M. L. M., Paul P., and Bakke O. (2011) Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat. Rev. Immunol. 11, 823–836 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
