

Sperm quality and quantity evolve through different selective processes in the Phasianidae
- PMID: 31848414
- PMCID: PMC6917726
- DOI: 10.1038/s41598-019-55822-3
Sperm quality and quantity evolve through different selective processes in the Phasianidae
Abstract
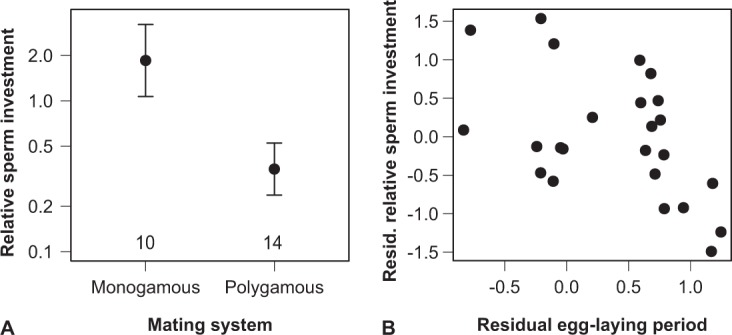
Sperm competition is often considered the primary selective force underlying the rapid and diversifying evolution of ejaculate traits. Yet, several recent studies have drawn attention to other forms of selection with the potential of exceeding the effects of sperm competition. Since ejaculates are complex, multivariate traits, it seems plausible that different ejaculate components vary in their responses to different selective pressures. Such information, however, is generally lacking as individual ejaculate traits tend to be studied in isolation. Here, we studied the macroevolutionary patterns of ejaculate volume, sperm number, sperm length and the proportion of viable normal sperm in response to varying levels of sperm competition, body size and the duration of female sperm storage in pheasants and allies (Phasianidae). Ejaculate volume, sperm number and sperm viability were all relatively higher in polygamous than in monogamous mating systems. However, whereas ejaculate volume additionally covaried with body size, sperm number instead increased with the female sperm-storage duration, in conjunction with a decrease in sperm length. Overall, our results revealed important details on how different forms of selection can jointly shape ejaculates as complex, composite traits.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Birkhead TR, Møller AP. Sexual selection and the temporal separation of reproductive events: sperm storage data from reptiles, birds and mammals. Biol. J. Linn. Soc. 1993;50:295–311. doi: 10.1111/j.1095-8312.1993.tb00933.x. - DOI
-
- Parker GA. Sperm competition and its evolutionary consequences in the insects. Biol. Rev. 1970;45:526–567. doi: 10.1111/j.1469-185X.1970.tb01176.x. - DOI
-
- Eberhard, W. G. Female Control: Sexual Selection by Cryptic Female Choice. (Princeton University Press, 1996).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
