

c-Abl Deficiency Provides Synaptic Resiliency Against Aβ-Oligomers
- PMID: 31849613
- PMCID: PMC6902026
- DOI: 10.3389/fncel.2019.00526
c-Abl Deficiency Provides Synaptic Resiliency Against Aβ-Oligomers
Abstract
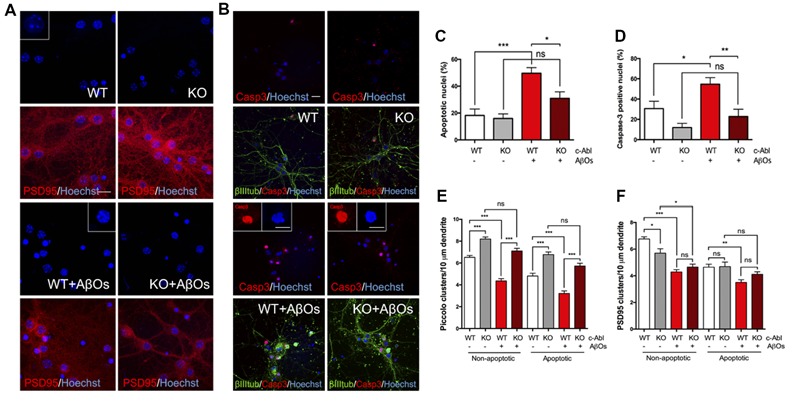
Spine pathology has been implicated in the early onset of Alzheimer's disease (AD), where Aβ-Oligomers (AβOs) cause synaptic dysfunction and loss. Previously, we described that pharmacological inhibition of c-Abl prevents AβOs-induced synaptic alterations. Hence, this kinase seems to be a key element in AD progression. Here, we studied the role of c-Abl on dendritic spine morphological changes induced by AβOs using c-Abl null neurons (c-Abl-KO). First, we characterized the effect of c-Abl deficiency on dendritic spine density and found that its absence increases dendritic spine density. While AβOs-treatment reduces the spine number in both wild-type (WT) and c-Abl-KO neurons, AβOs-driven spine density loss was not affected by c-Abl. We then characterized AβOs-induced morphological changes in dendritic spines of c-Abl-KO neurons. AβOs induced a decrease in the number of mushroom spines in c-Abl-KO neurons while preserving the populations of immature stubby, thin, and filopodia spines. Furthermore, synaptic contacts evaluated by PSD95/Piccolo clustering and cell viability were preserved in AβOs-exposed c-Abl-KO neurons. In conclusion, our results indicate that in the presence of AβOs c-Abl participates in synaptic contact removal, increasing susceptibility to AβOs damage. Its deficiency increases the immature spine population reducing AβOs-induced synapse elimination. Therefore, c-Abl signaling could be a relevant actor in the early stages of AD.
Keywords: Alzheimer’s disease; Aβ-oligomers; c-Abl tyrosine kinase; dendritic spines; synapse.
Copyright © 2019 Gutierrez, Vargas, Chandia-Cristi, de la Fuente, Leal and Alvarez.
Figures
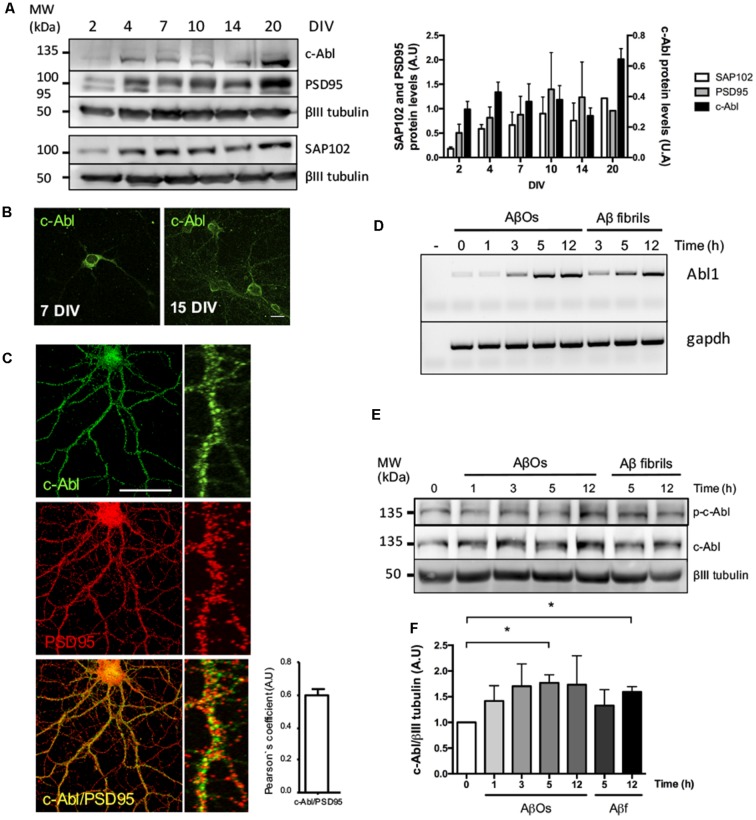
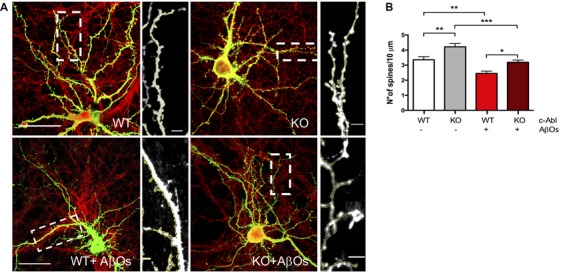
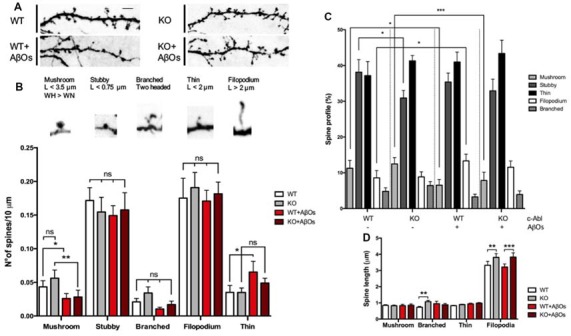
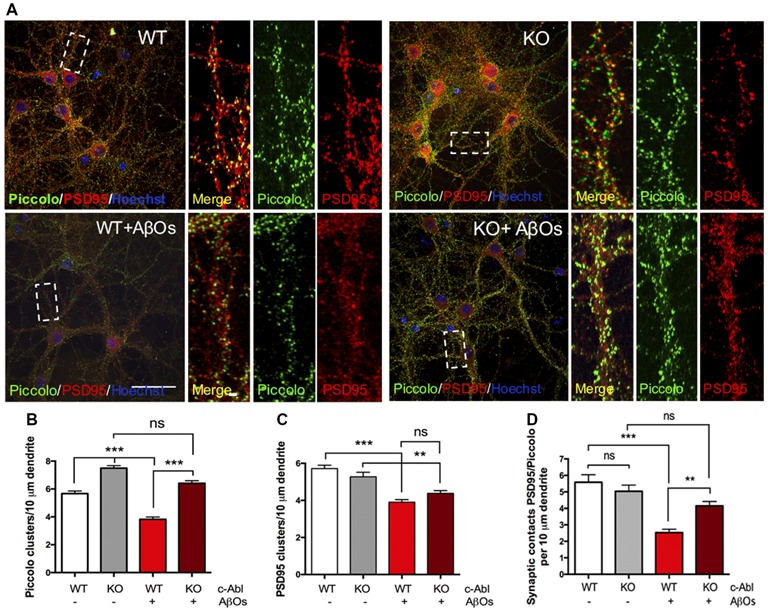
References
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous