

Snake Venom Cathelicidins as Natural Antimicrobial Peptides
- PMID: 31849667
- PMCID: PMC6895205
- DOI: 10.3389/fphar.2019.01415
Snake Venom Cathelicidins as Natural Antimicrobial Peptides
Abstract
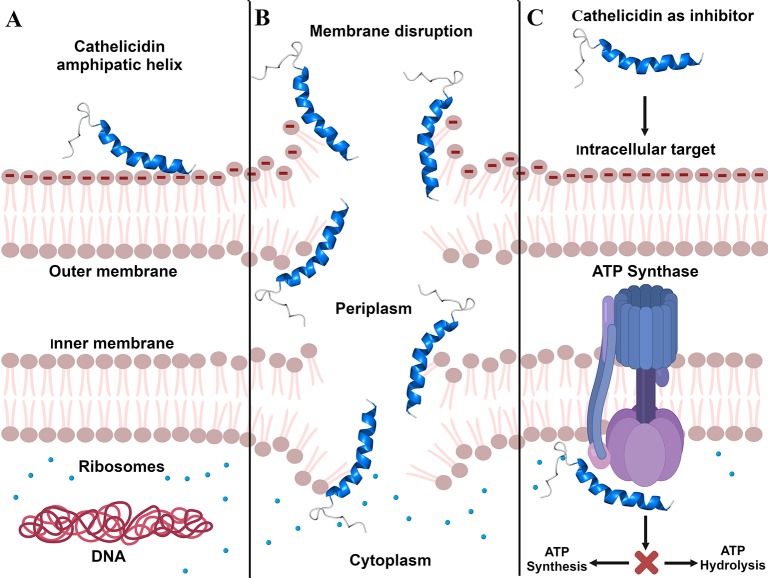
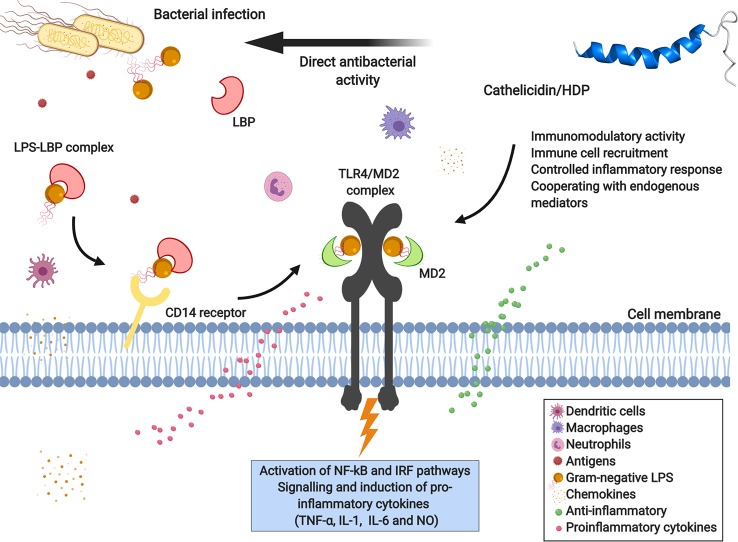
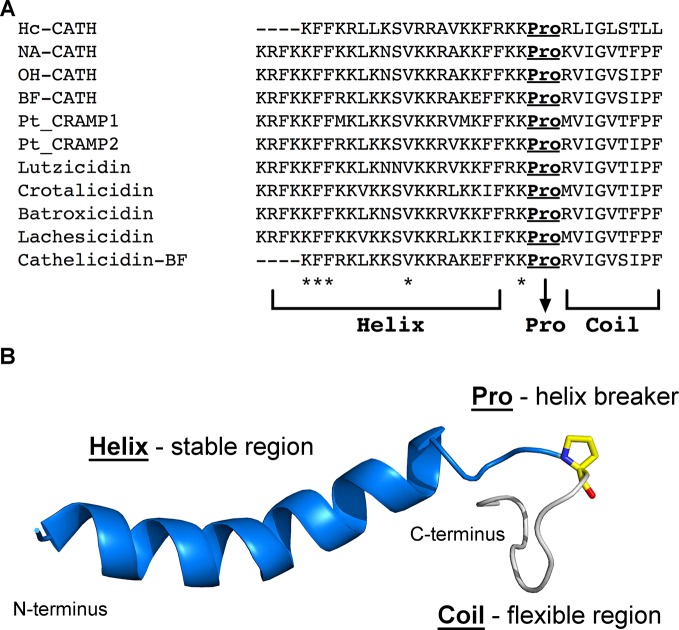
Bioactive small molecules isolated from animals, plants, fungi and bacteria, including natural antimicrobial peptides, have shown great therapeutic potential worldwide. Among these peptides, snake venom cathelicidins are being widely exploited, because the variation in the composition of the venom reflects a range of biological activities that may be of biotechnological interest. Cathelicidins are short, cationic, and amphipathic molecules. They play an important role in host defense against microbial infections. We are currently facing a strong limitation on pharmacological interventions for infection control, which has become increasingly complex due to the lack of effective therapeutic options. In this review, we will focus on natural snake venom cathelicidins as promising candidates for the development of new antibacterial agents to fight antibiotic-resistant bacteria. We will highlight their antibacterial and antibiofilm activities, mechanism of action, and modulation of the innate immune response.
Keywords: antimicrobial peptides; cathelicidins; host-defense peptides; natural peptides; snake venom.
Copyright © 2019 de Barros, Gonçalves, Cardoso, Santos, Franco and Cândido.
Figures
References
-
- Almeida J. R., Mendes B., Lancellotti M., Marangoni S., Vale N., Passos Ó., et al. (2018). A novel synthetic peptide inspired on Lys49 phospholipase A2 from Crotalus oreganus abyssus snake venom active against multidrug-resistant clinical isolates. Eur. J. Med. Chem. 149, 248–256. 10.1016/j.ejmech.2018.02.055 - DOI - PubMed
Publication types
LinkOut - more resources
Full Text Sources