

Topologically Associated Domains Delineate Susceptibility to Somatic Hypermutation
- PMID: 31851922
- PMCID: PMC6980758
- DOI: 10.1016/j.celrep.2019.11.039
Topologically Associated Domains Delineate Susceptibility to Somatic Hypermutation
Abstract
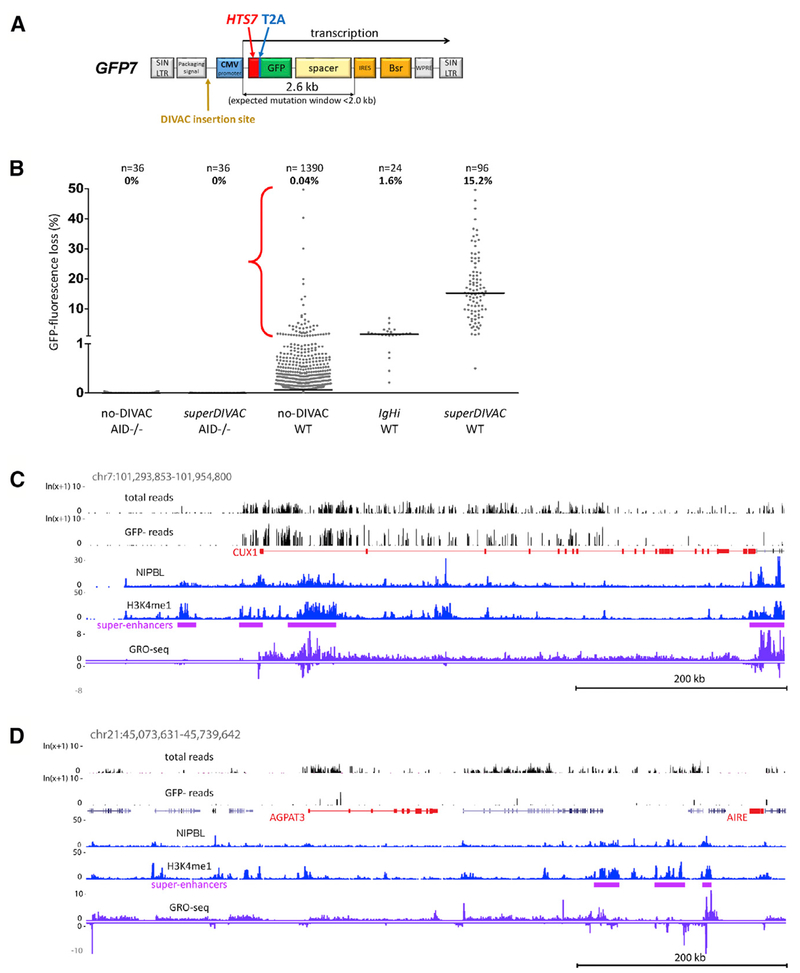
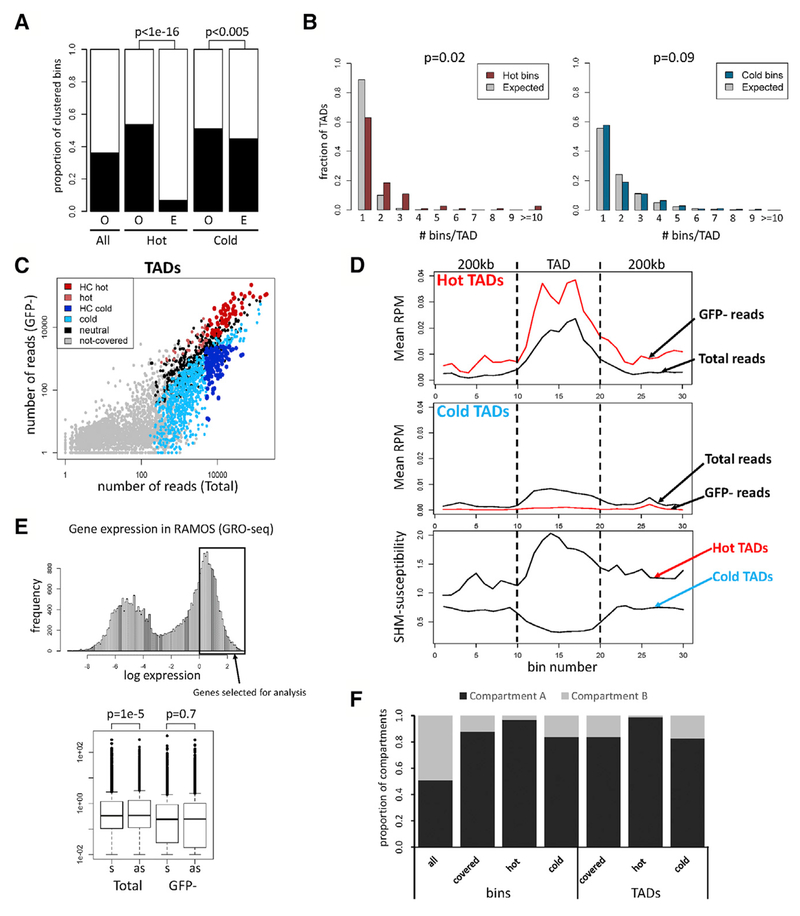
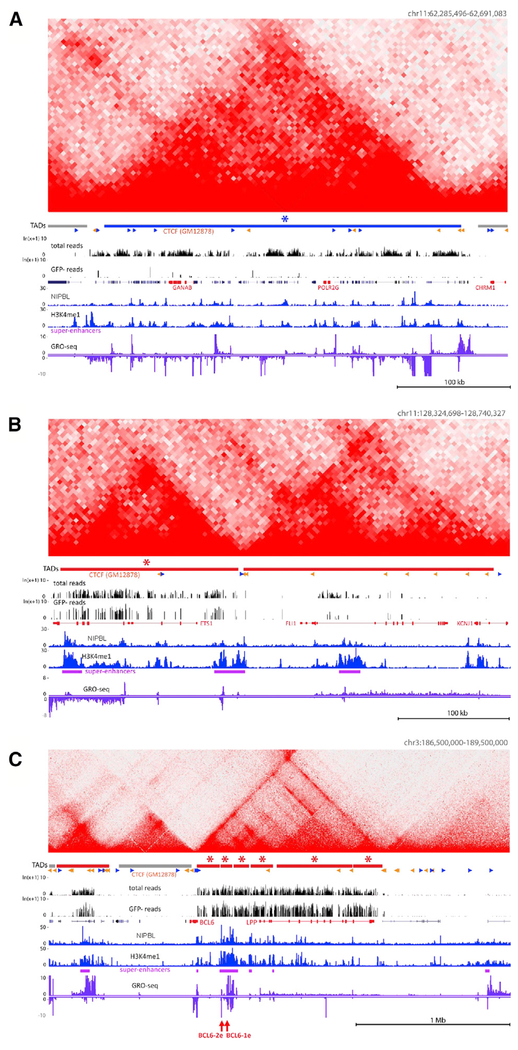
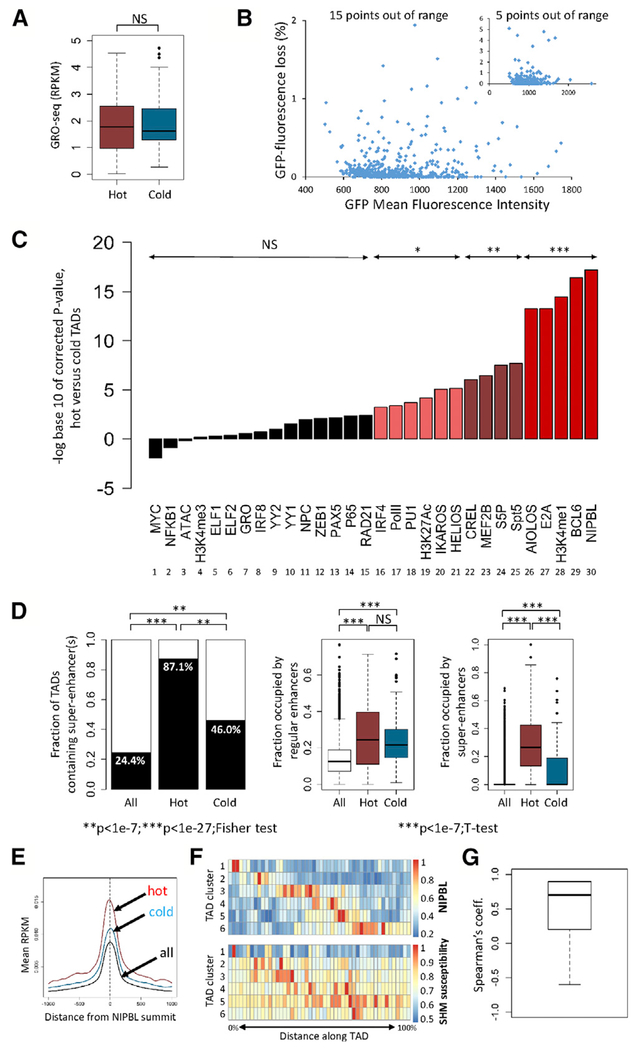
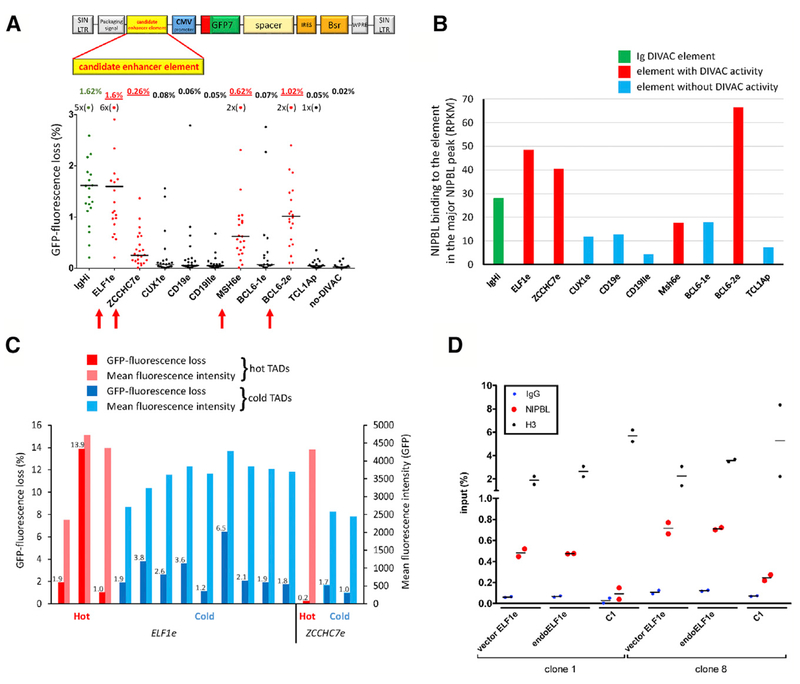
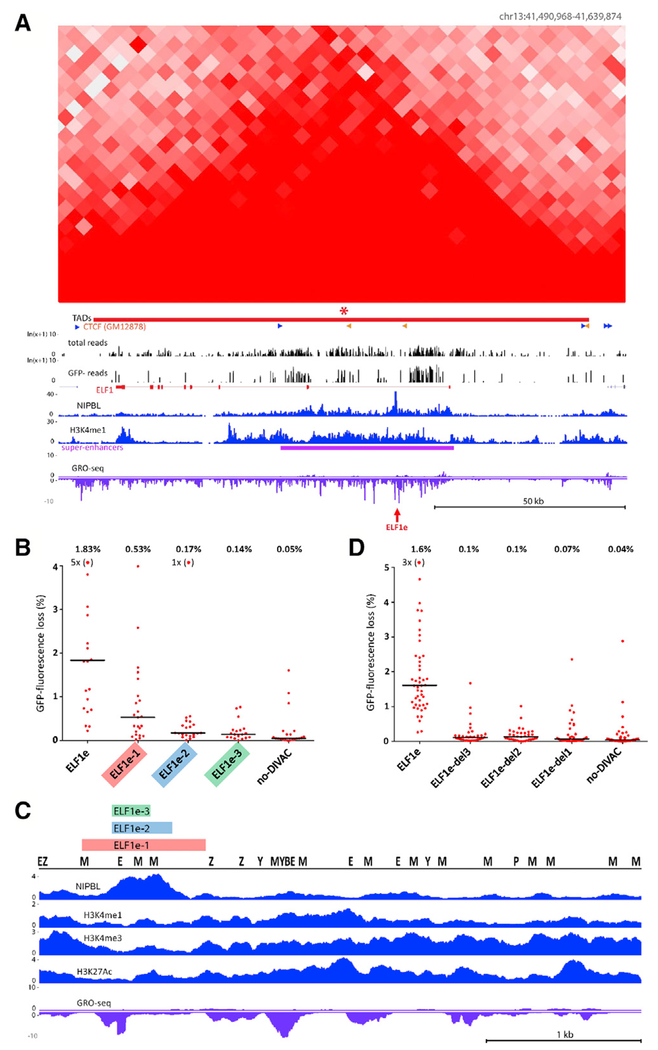
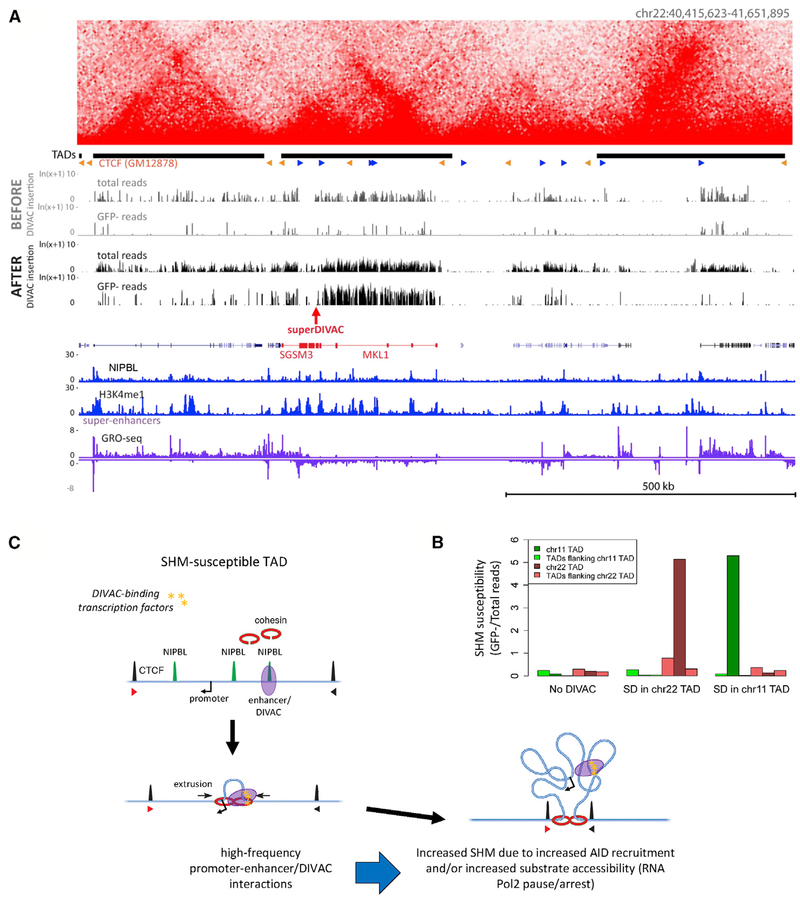
Somatic hypermutation (SHM) introduces point mutations into immunoglobulin (Ig) genes but also causes mutations in other parts of the genome. We have used lentiviral SHM reporter vectors to identify regions of the genome that are susceptible ("hot") and resistant ("cold") to SHM, revealing that SHM susceptibility and resistance are often properties of entire topologically associated domains (TADs). Comparison of hot and cold TADs reveals that while levels of transcription are equivalent, hot TADs are enriched for the cohesin loader NIPBL, super-enhancers, markers of paused/stalled RNA polymerase 2, and multiple important B cell transcription factors. We demonstrate that at least some hot TADs contain enhancers that possess SHM targeting activity and that insertion of a strong Ig SHM-targeting element into a cold TAD renders it hot. Our findings lead to a model for SHM susceptibility involving the cooperative action of cis-acting SHM targeting elements and the dynamic and architectural properties of TADs.
Keywords: activation induced deaminase; chromatin loop extrusion; chromatin structure; somatic hypermutation; topologically associated domain; transcription factor.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
References
-
- Basso K, and Dalla-Favera R (2012). Roles of BCL6 in normal and transformed germinal center B cells. Immunol. Rev 247, 172–183. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
