

Spatial Genome Re-organization between Fetal and Adult Hematopoietic Stem Cells
- PMID: 31851943
- PMCID: PMC7262670
- DOI: 10.1016/j.celrep.2019.11.065
Spatial Genome Re-organization between Fetal and Adult Hematopoietic Stem Cells
Abstract
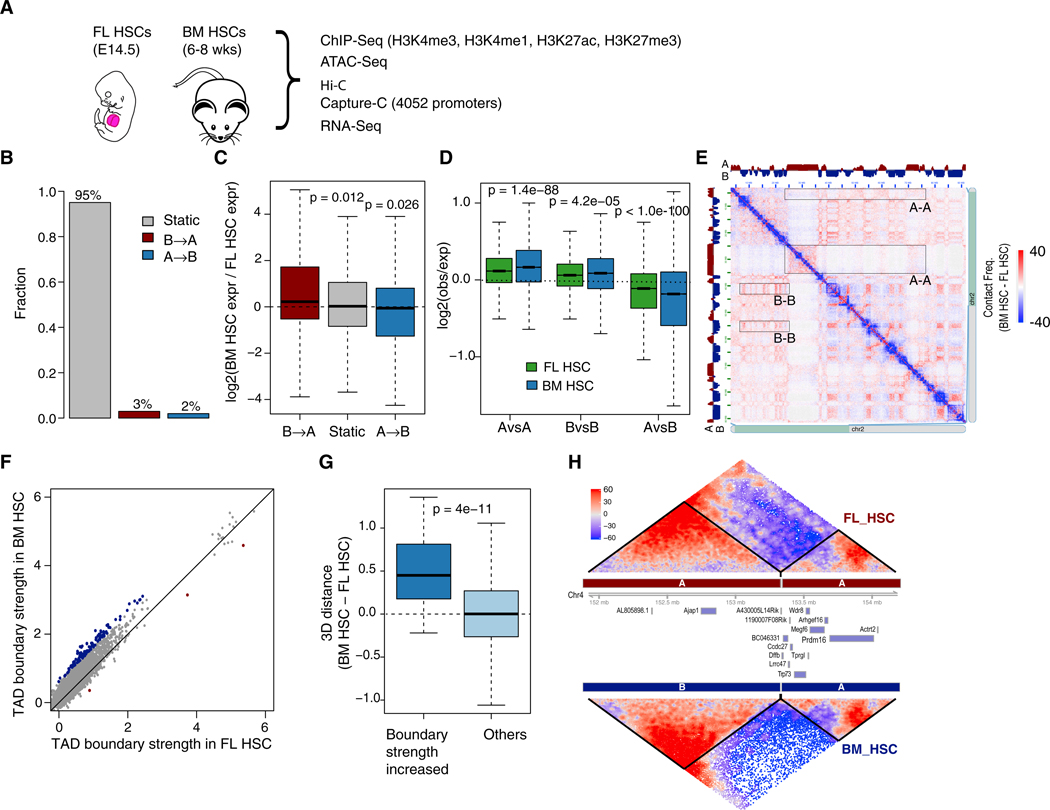
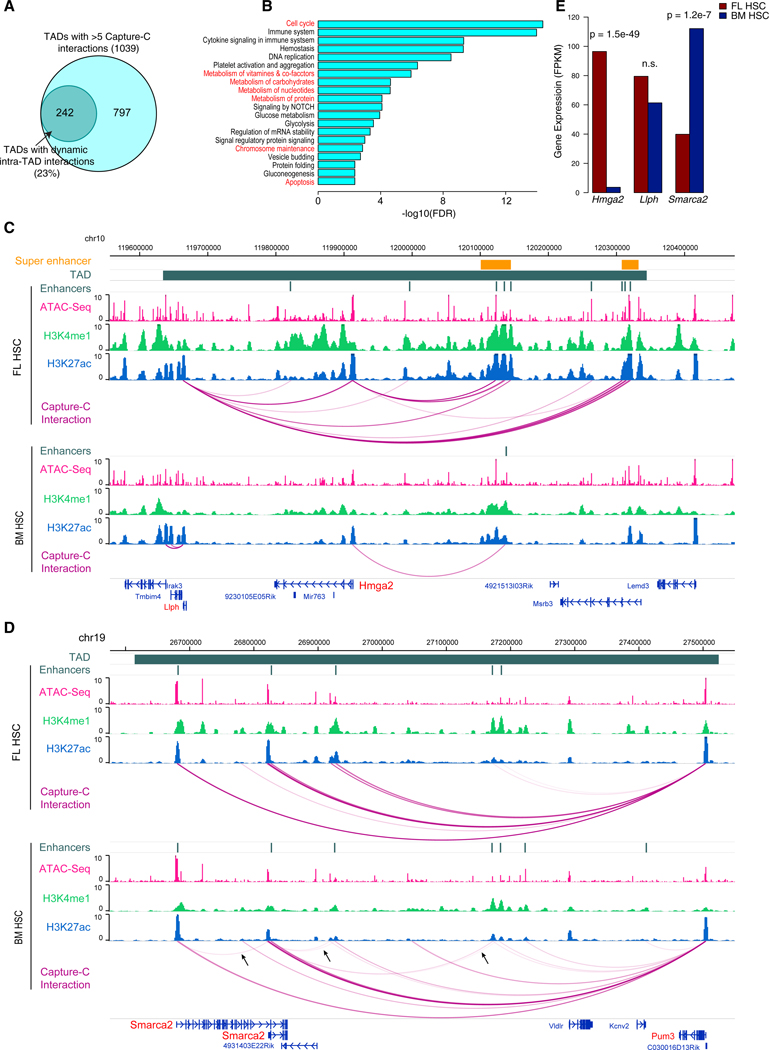
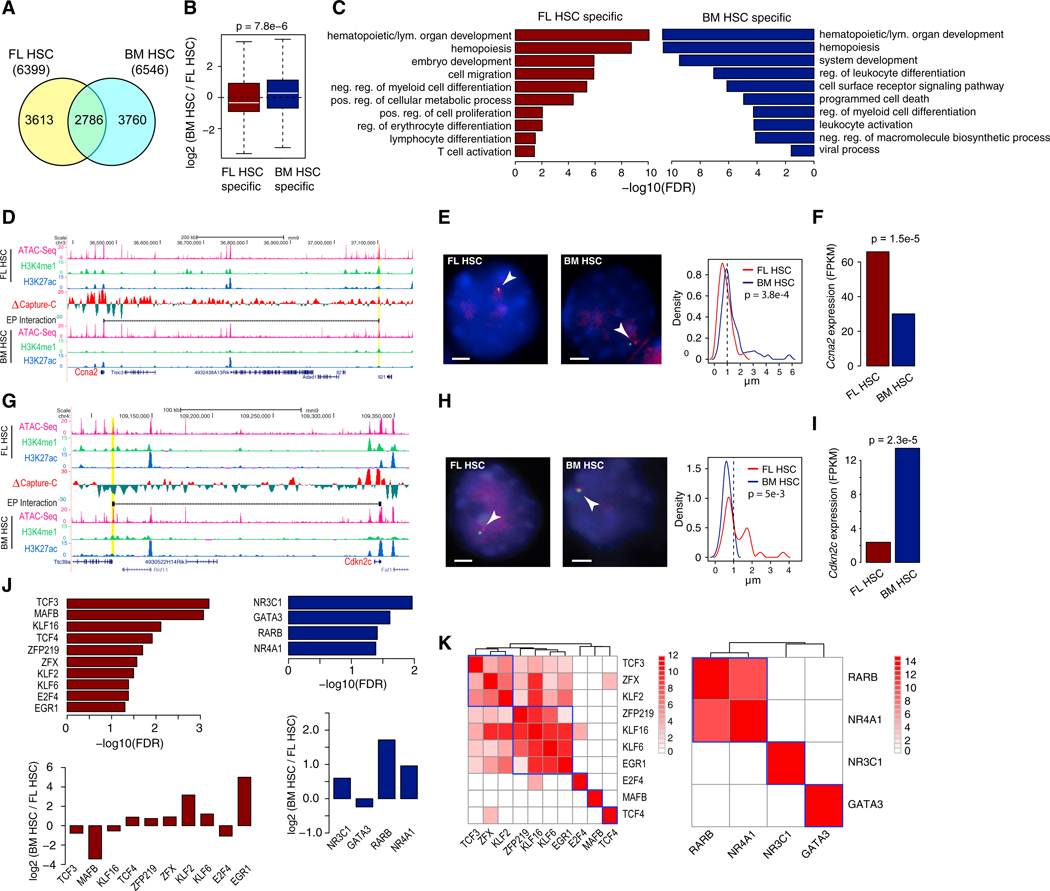
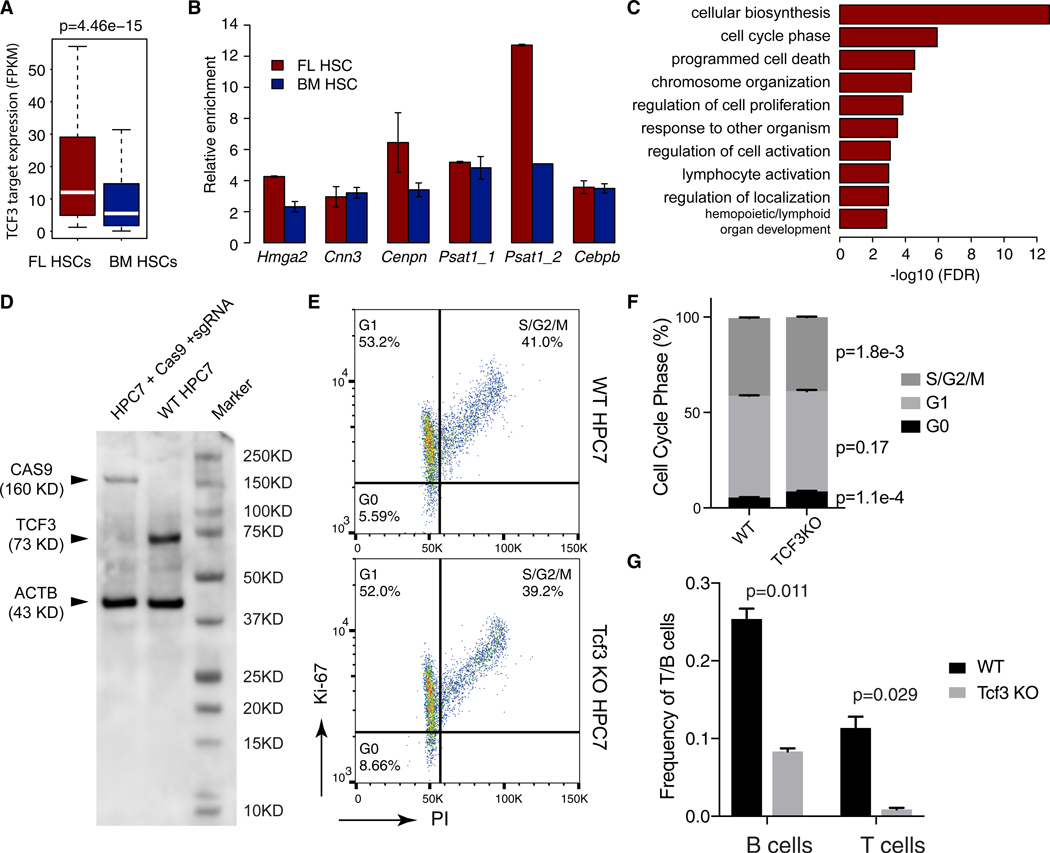
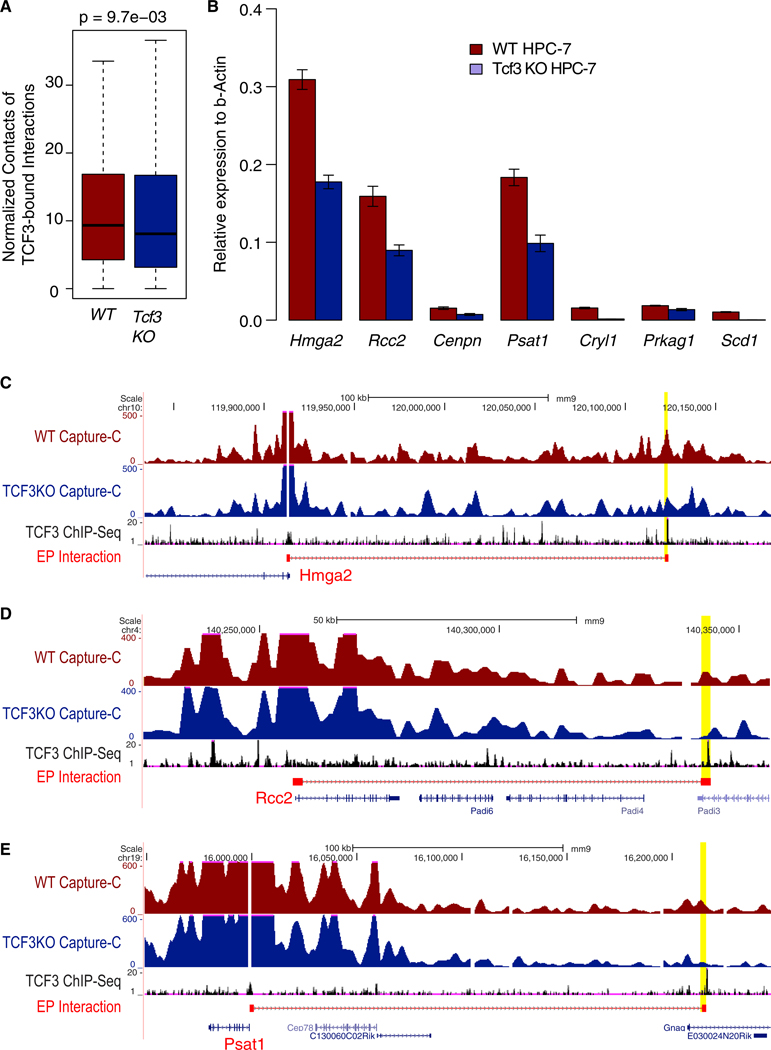
Fetal hematopoietic stem cells (HSCs) undergo a developmental switch to become adult HSCs with distinct functional properties. To better understand the molecular mechanisms underlying the developmental switch, we have conducted deep sequencing of the 3D genome, epigenome, and transcriptome of fetal and adult HSCs in mouse. We find that chromosomal compartments and topologically associating domains (TADs) are largely conserved between fetal and adult HSCs. However, there is a global trend of increased compartmentalization and TAD boundary strength in adult HSCs. In contrast, intra-TAD chromatin interactions are much more dynamic and widespread, involving over a thousand gene promoters and distal enhancers. These developmental-stage-specific enhancer-promoter interactions are mediated by different sets of transcription factors, such as TCF3 and MAFB in fetal HSCs, versus NR4A1 and GATA3 in adult HSCs. Loss-of-function studies of TCF3 confirm the role of TCF3 in mediating condition-specific enhancer-promoter interactions and gene regulation in fetal HSCs.
Keywords: 3D genome; enhancer-promoter interaction; epigenomics; hematopoiesis; transcriptome.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Adolfsson J, Borge OJ, Bryder D, Theilgaard-Mönch K, Astrand-Grund-ström I, Sitnicka E, Sasaki Y, and Jacobsen SE (2001). Upregulation of Flt3 expression within the bone marrow Lin(−)Sca1(+)c-kit(+) stem cell compartment is accompanied by loss of self-renewal capacity. Immunity 15, 659–669. - PubMed
-
- Benz C, Copley MR, Kent DG, Wohrer S, Cortes A, Aghaeepour N, Ma E, Mader H, Rowe K, Day C, et al. (2012). Hematopoietic stem cell subtypes expand differentially during development and display distinct lymphopoietic programs. Cell Stem Cell 10, 273–283. - PubMed
-
- Bonev B, and Cavalli G. (2016). Organization and function of the 3D genome. Nat. Rev. Genet 17, 661–678. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
