

Diatom Molecular Research Comes of Age: Model Species for Studying Phytoplankton Biology and Diversity
- PMID: 31852772
- PMCID: PMC7054031
- DOI: 10.1105/tpc.19.00158
Diatom Molecular Research Comes of Age: Model Species for Studying Phytoplankton Biology and Diversity
Abstract
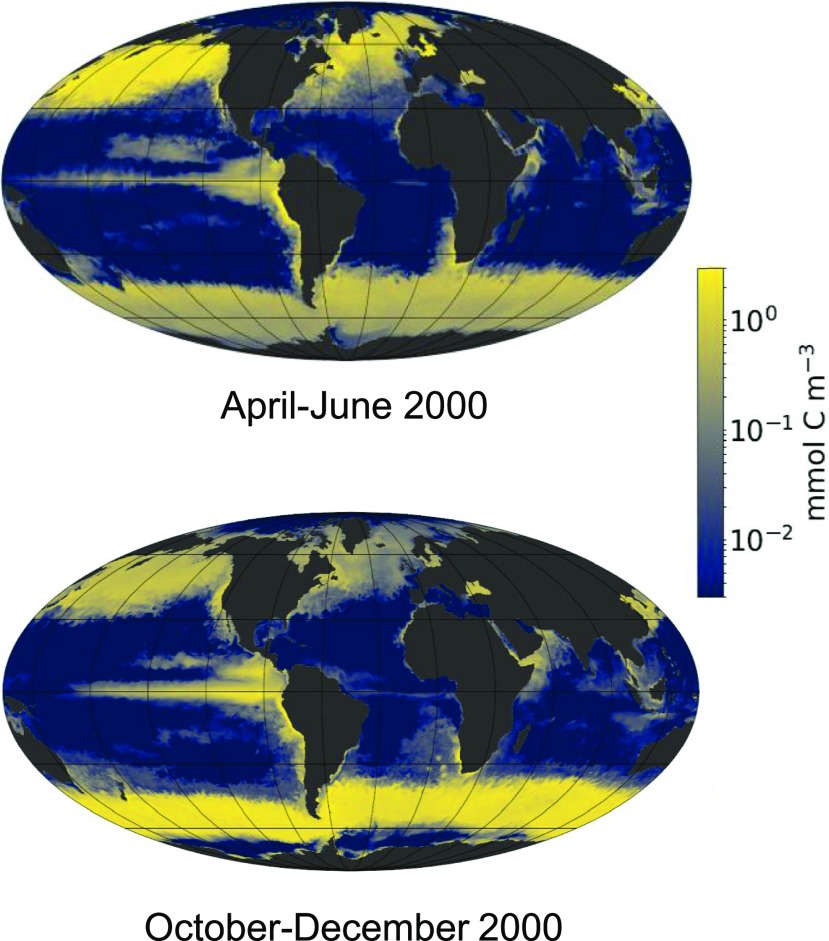
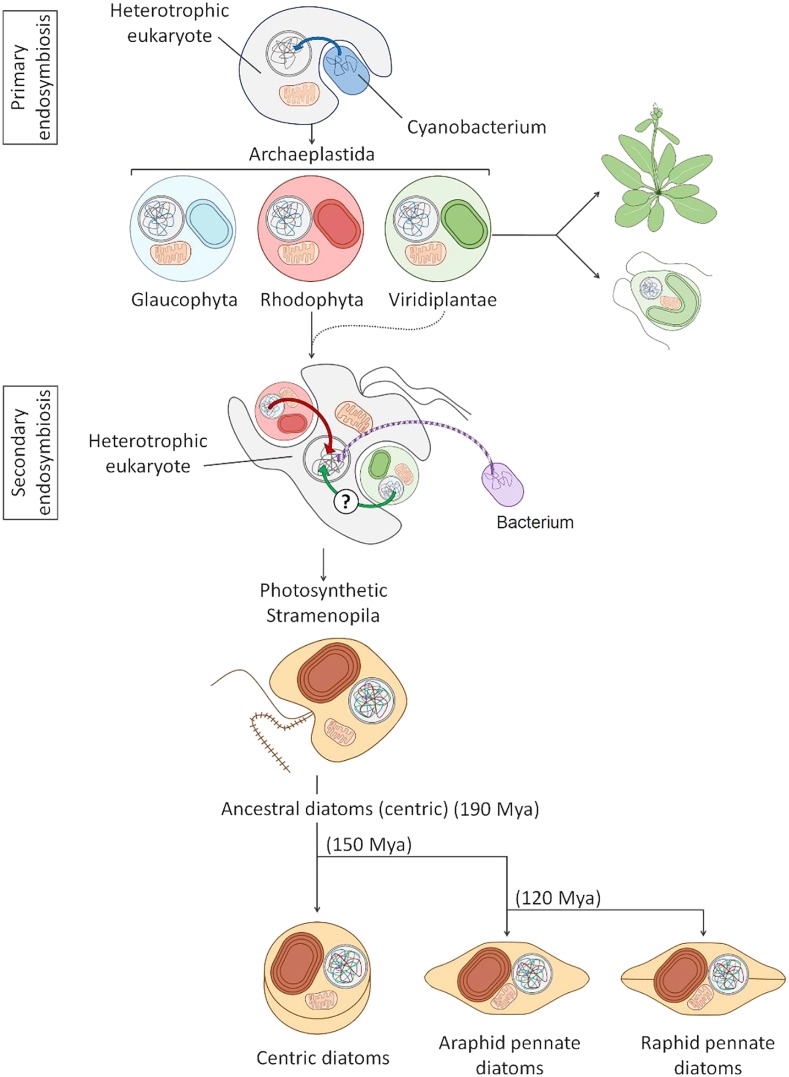
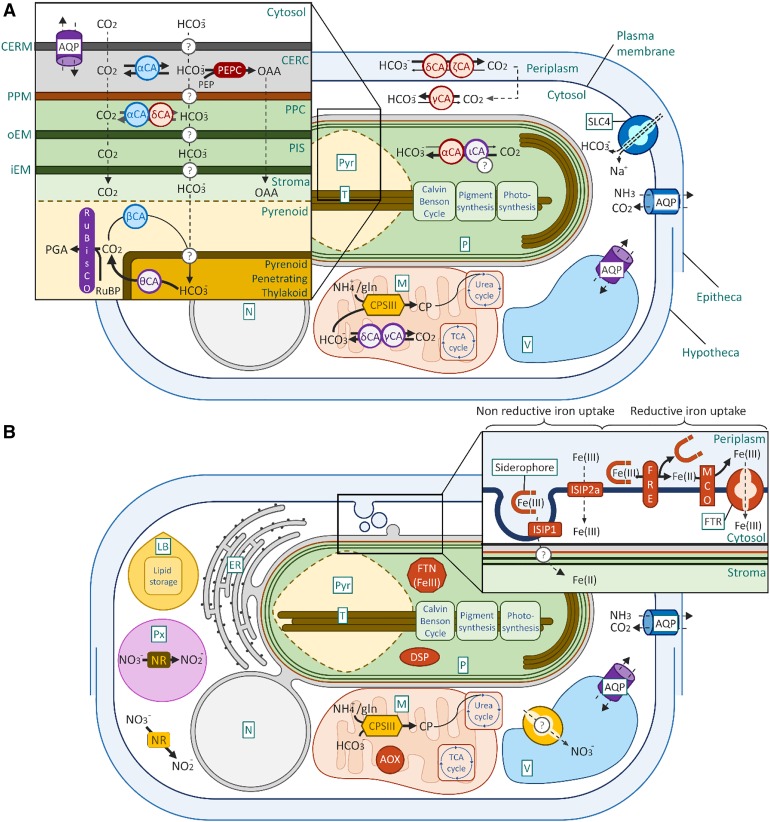
Diatoms are the world's most diverse group of algae, comprising at least 100,000 species. Contributing ∼20% of annual global carbon fixation, they underpin major aquatic food webs and drive global biogeochemical cycles. Over the past two decades, Thalassiosira pseudonana and Phaeodactylum tricornutum have become the most important model systems for diatom molecular research, ranging from cell biology to ecophysiology, due to their rapid growth rates, small genomes, and the cumulative wealth of associated genetic resources. To explore the evolutionary divergence of diatoms, additional model species are emerging, such as Fragilariopsis cylindrus and Pseudo-nitzschia multistriata Here, we describe how functional genomics and reverse genetics have contributed to our understanding of this important class of microalgae in the context of evolution, cell biology, and metabolic adaptations. Our review will also highlight promising areas of investigation into the diversity of these photosynthetic organisms, including the discovery of new molecular pathways governing the life of secondary plastid-bearing organisms in aquatic environments.
© 2020 American Society of Plant Biologists. All rights reserved.
Figures

References
-
- Ahmed S., et al. (2014). A haploid system of sex determination in the brown alga Ectocarpus sp. Curr. Biol. 24: 1945–1957. - PubMed
-
- Allen A.E., Dupont C.L., Oborník M., Horák A., Nunes-Nesi A., McCrow J.P., Zheng H., Johnson D.A., Hu H., Fernie A.R., Bowler C. (2011). Evolution and metabolic significance of the urea cycle in photosynthetic diatoms. Nature 473: 203–207. - PubMed
-
- Allen J. (2002). Photosynthesis of ATP-electrons, proton pumps, rotors, and poise. Cell 110: 273–276. - PubMed
-
- Amato A., Orsini L. (2015). Rare interspecific breeding in Pseudo-nitzschia (Bacillariophyceae). Phytotaxa 217: 145–154.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
