

Human genetics and neuropathology suggest a link between miR-218 and amyotrophic lateral sclerosis pathophysiology
- PMID: 31852800
- PMCID: PMC7057809
- DOI: 10.1126/scitranslmed.aav5264
Human genetics and neuropathology suggest a link between miR-218 and amyotrophic lateral sclerosis pathophysiology
Abstract
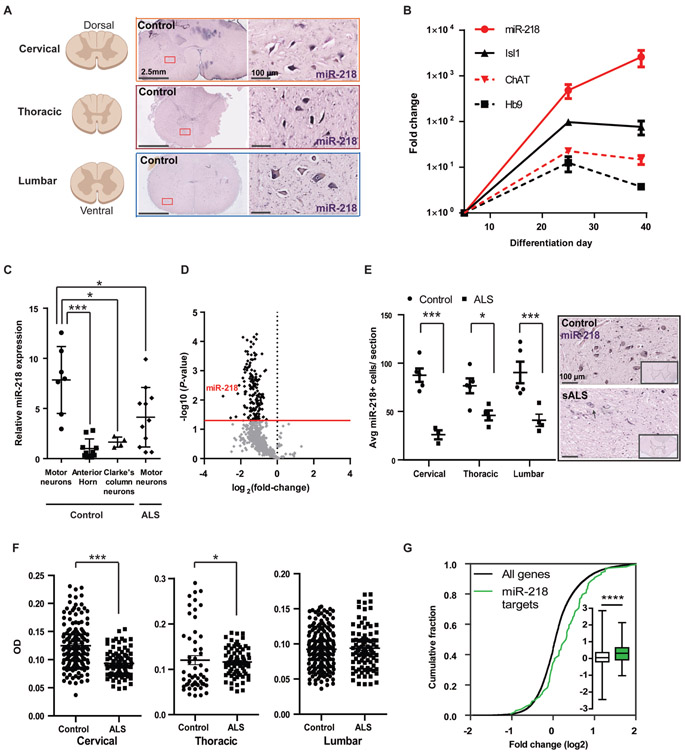
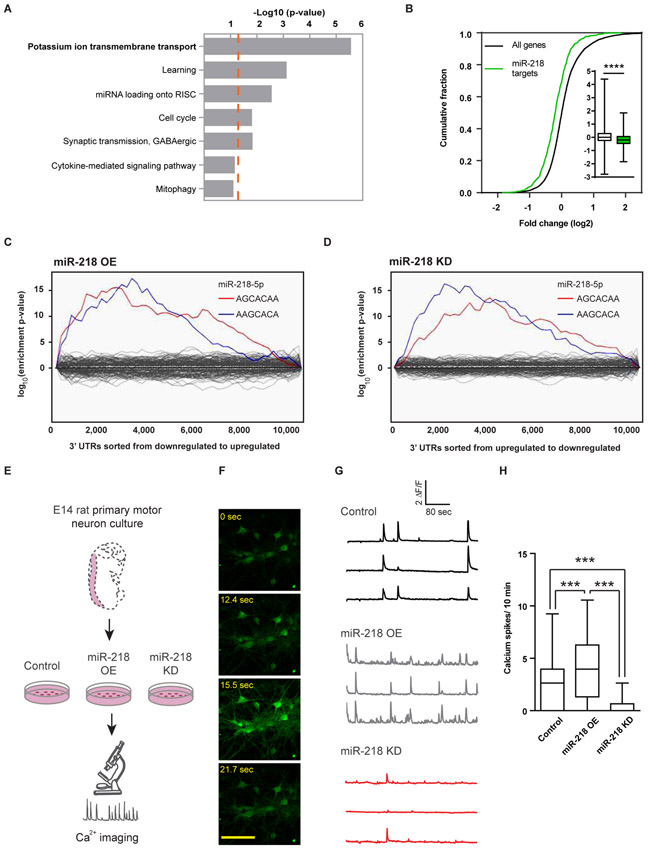
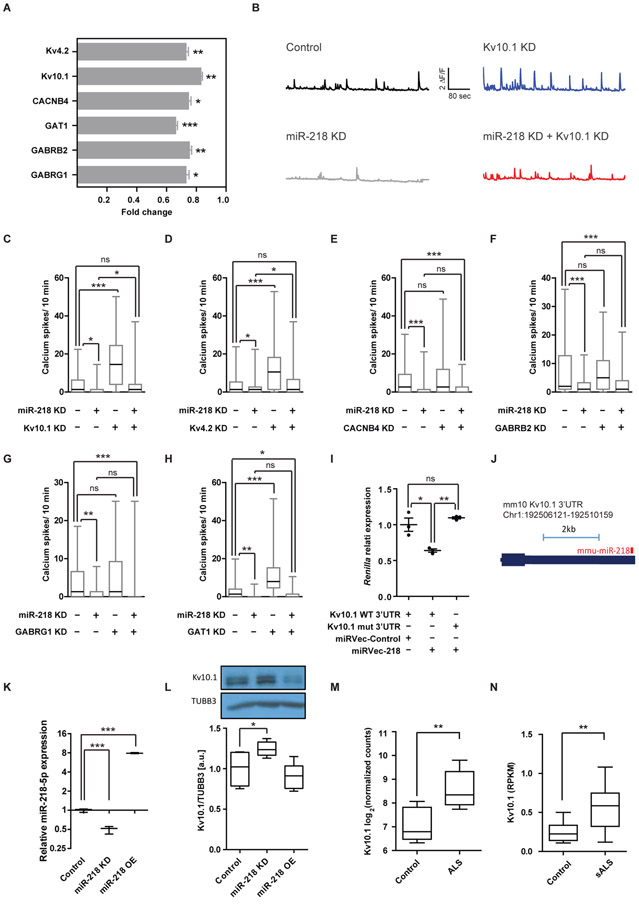
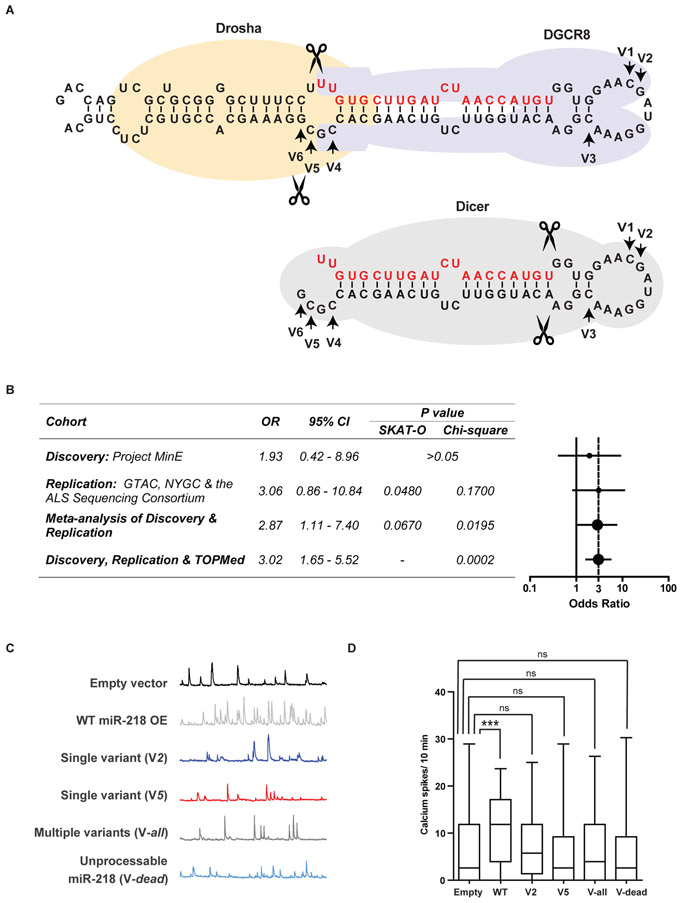
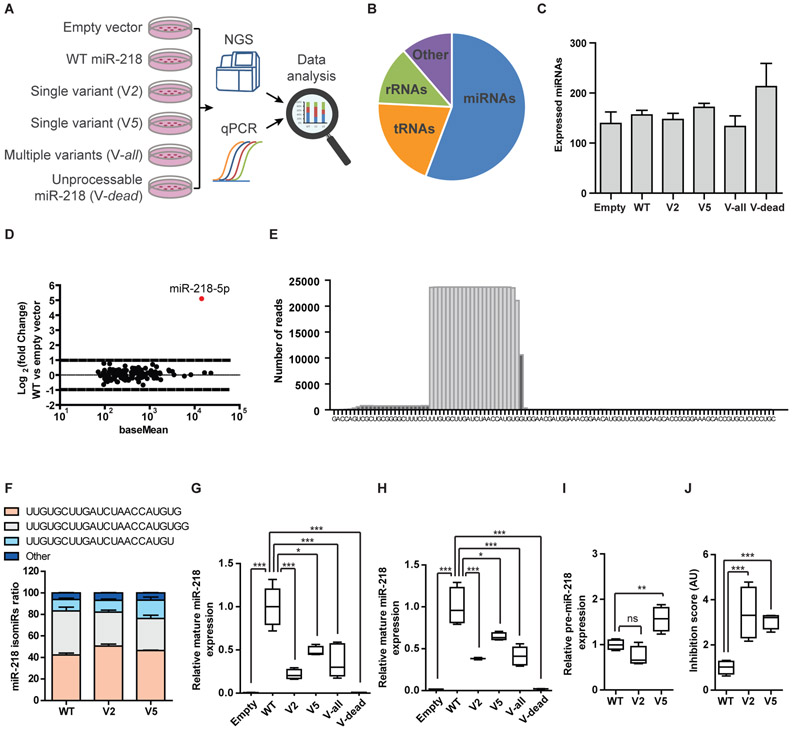
Motor neuron-specific microRNA-218 (miR-218) has recently received attention because of its roles in mouse development. However, miR-218 relevance to human motor neuron disease was not yet explored. Here, we demonstrate by neuropathology that miR-218 is abundant in healthy human motor neurons. However, in amyotrophic lateral sclerosis (ALS) motor neurons, miR-218 is down-regulated and its mRNA targets are reciprocally up-regulated (derepressed). We further identify the potassium channel Kv10.1 as a new miR-218 direct target that controls neuronal activity. In addition, we screened thousands of ALS genomes and identified six rare variants in the human miR-218-2 sequence. miR-218 gene variants fail to regulate neuron activity, suggesting the importance of this small endogenous RNA for neuronal robustness. The underlying mechanisms involve inhibition of miR-218 biogenesis and reduced processing by DICER. Therefore, miR-218 activity in motor neurons may be susceptible to failure in human ALS, suggesting that miR-218 may be a potential therapeutic target in motor neuron disease.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
References
-
- Emde A, Eitan C, Liou LL, Libby RT, Rivkin N, Magen I, Reichenstein I, Oppenheim H, Eilam R, Silvestroni A, Alajajian B, Ben-Dov IZ, Aebischer J, Savidor A, Levin Y, Sons R, Hammond SM, Ravits JM, Moller T, Hornstein E, Dysregulated miRNA biogenesis downstream of cellular stress and ALS-causing mutations: a new mechanism for ALS. The EMBO journal 34, 2633–2651 (2015). - PMC - PubMed
-
- Hoye ML, Koval ED, Wegener AJ, Hyman TS, Yang C, O'Brien DR, Miller RL, Cole T, Schoch KM, Shen T, Kunikata T, Richard JP, Gutmann DH, Maragakis NJ, Kordasiewicz HB, Dougherty JD, Miller TM, MicroRNA Profiling Reveals Marker of Motor Neuron Disease in ALS Models. J Neurosci 37, 5574–5586 (2017). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- F31 NS092340/NS/NINDS NIH HHS/United States
- R01 NS092705/NS/NINDS NIH HHS/United States
- R56 NS073873/NS/NINDS NIH HHS/United States
- MR/L501529/1/MRC_/Medical Research Council/United Kingdom
- G0600974/MRC_/Medical Research Council/United Kingdom
- R01 NS111990/NS/NINDS NIH HHS/United States
- R01 NS073873/NS/NINDS NIH HHS/United States
- R01 NS078398/NS/NINDS NIH HHS/United States
- TURNER/OCT15/972-797/MNDA_/Motor Neurone Disease Association/United Kingdom
- R01 NS104022/NS/NINDS NIH HHS/United States
- MR/R024804/1/MRC_/Medical Research Council/United Kingdom
- WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
