

Beyond the darkness: recent lessons from etiolation and de-etiolation studies
- PMID: 31854450
- PMCID: PMC7031072
- DOI: 10.1093/jxb/erz496
Beyond the darkness: recent lessons from etiolation and de-etiolation studies
Abstract
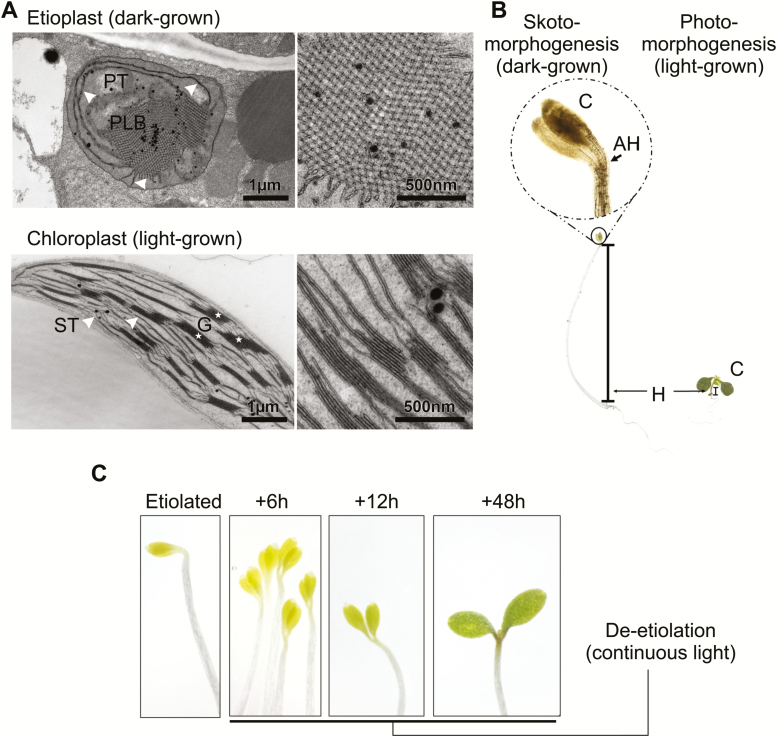
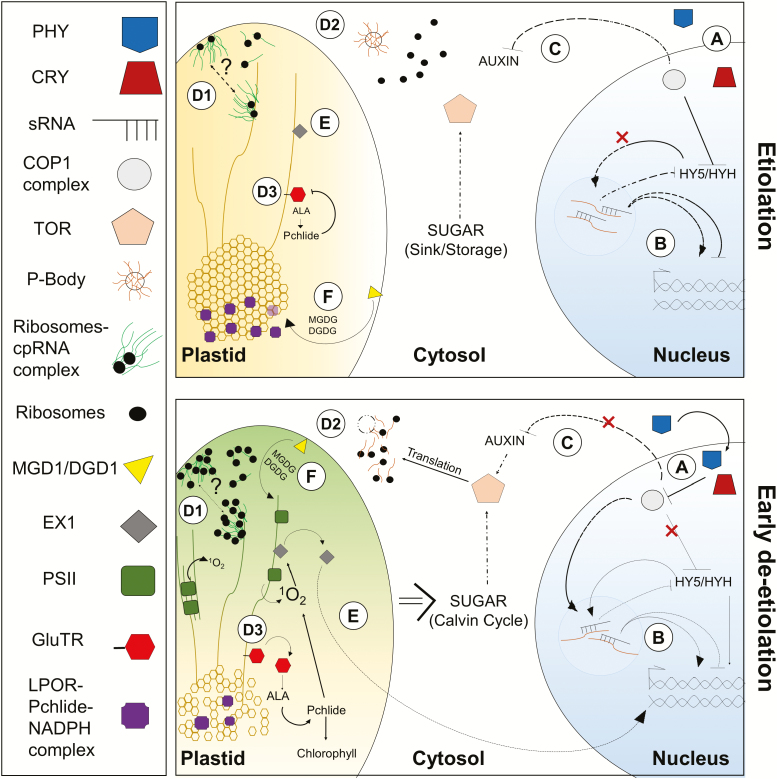
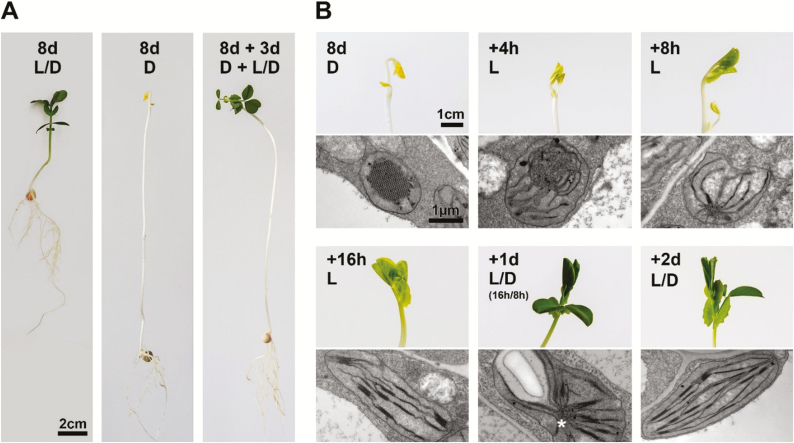
The state of etiolation is generally defined by the presence of non-green plastids (etioplasts) in plant tissues that would normally contain chloroplasts. In the commonly used dark-grown seedling system, etiolation is coupled with a type of growth called skotomorphogenesis. Upon illumination, de-etiolation occurs, marked by the transition from etioplast to chloroplast, and, at the seedling level, a switch to photomorphogenic growth. Etiolation and de-etiolation systems are therefore important for understanding both the acquisition of photosynthetic capacity during chloroplast biogenesis and plant responses to light-the most relevant signal in the life and growth of the organism. In this review, we discuss recent discoveries (within the past 2-3 years) in the field of etiolation and de-etiolation, with a particular focus on post-transcriptional processes and ultrastructural changes. We further discuss ambiguities in definitions of the term 'etiolation', and benefits and biases of common etiolation/de-etiolation systems. Finally, we raise several open questions and future research possibilities.
Keywords: chloroplast biogenesis; de-etiolation; etiolation; etioplast; prolamellar body; skotomorphogenesis.
© The Author(s) 2019. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Achkar NP, Cho SK, Poulsen C, et al. 2018. A quick HYL1-dependent reactivation of microRNA production is required for a proper developmental response after extended periods of light deprivation. Developmental Cell 46, 236–247.e6. - PubMed
-
- Aronsson H, Schöttler MA, Kelly AA, Sundqvist C, Dörmann P, Karim S, Jarvis P. 2008. Monogalactosyldiacylglycerol deficiency in Arabidopsis affects pigment composition in the prolamellar body and impairs thylakoid membrane energization and photoprotection in leaves. Plant Physiology 148, 580–592. - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
