

VDAC oligomers form mitochondrial pores to release mtDNA fragments and promote lupus-like disease
- PMID: 31857488
- PMCID: PMC8325171
- DOI: 10.1126/science.aav4011
VDAC oligomers form mitochondrial pores to release mtDNA fragments and promote lupus-like disease
Abstract
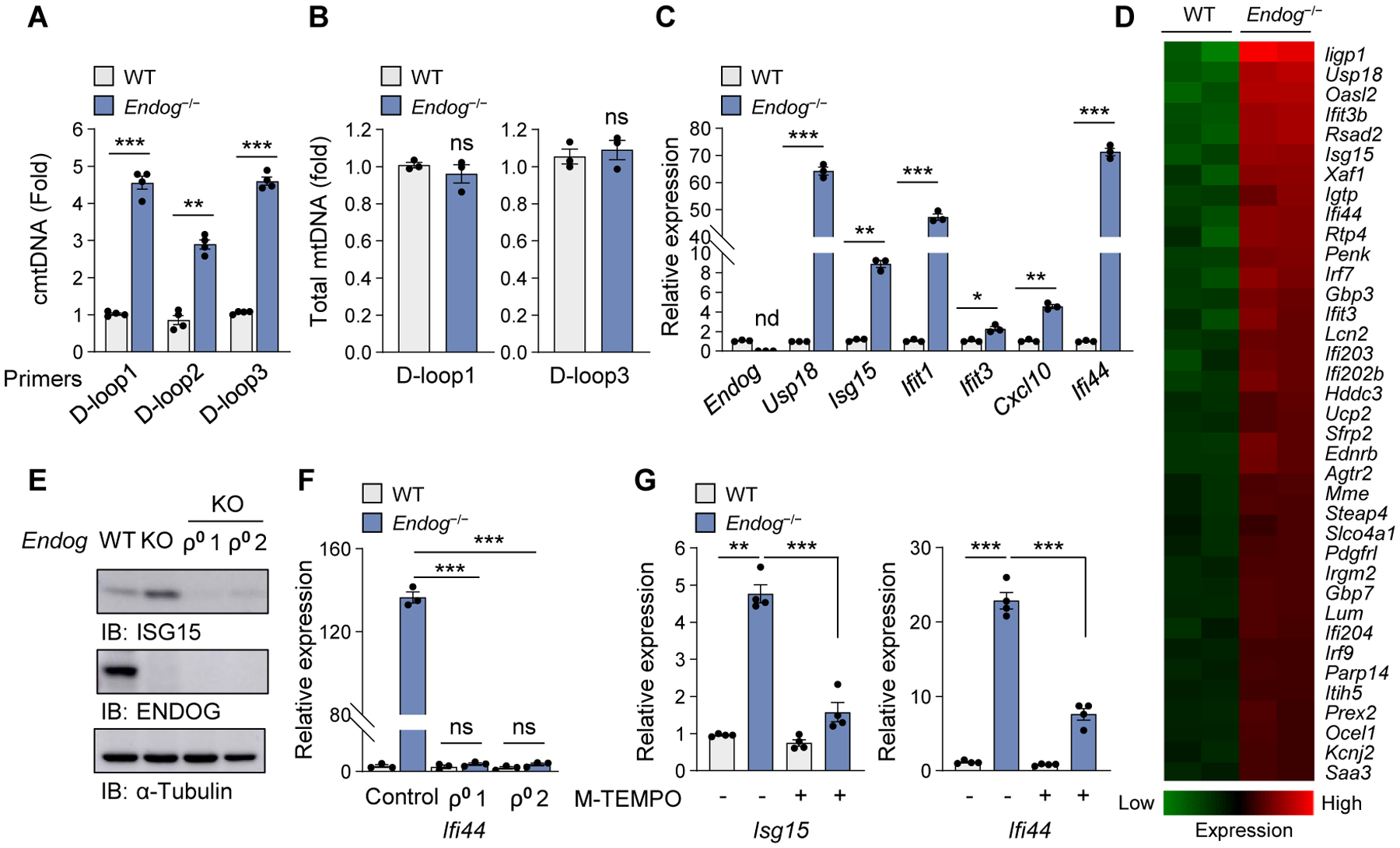
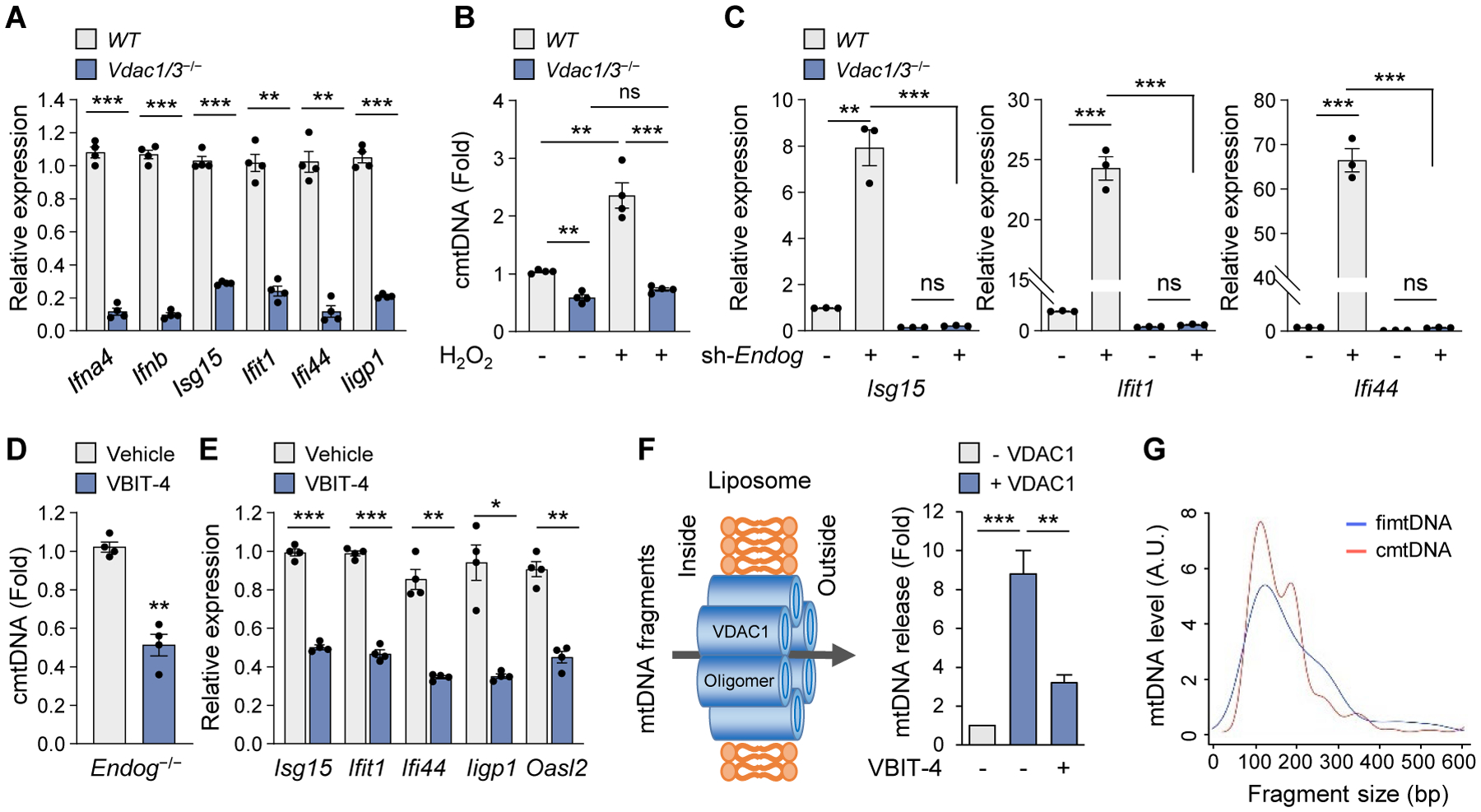
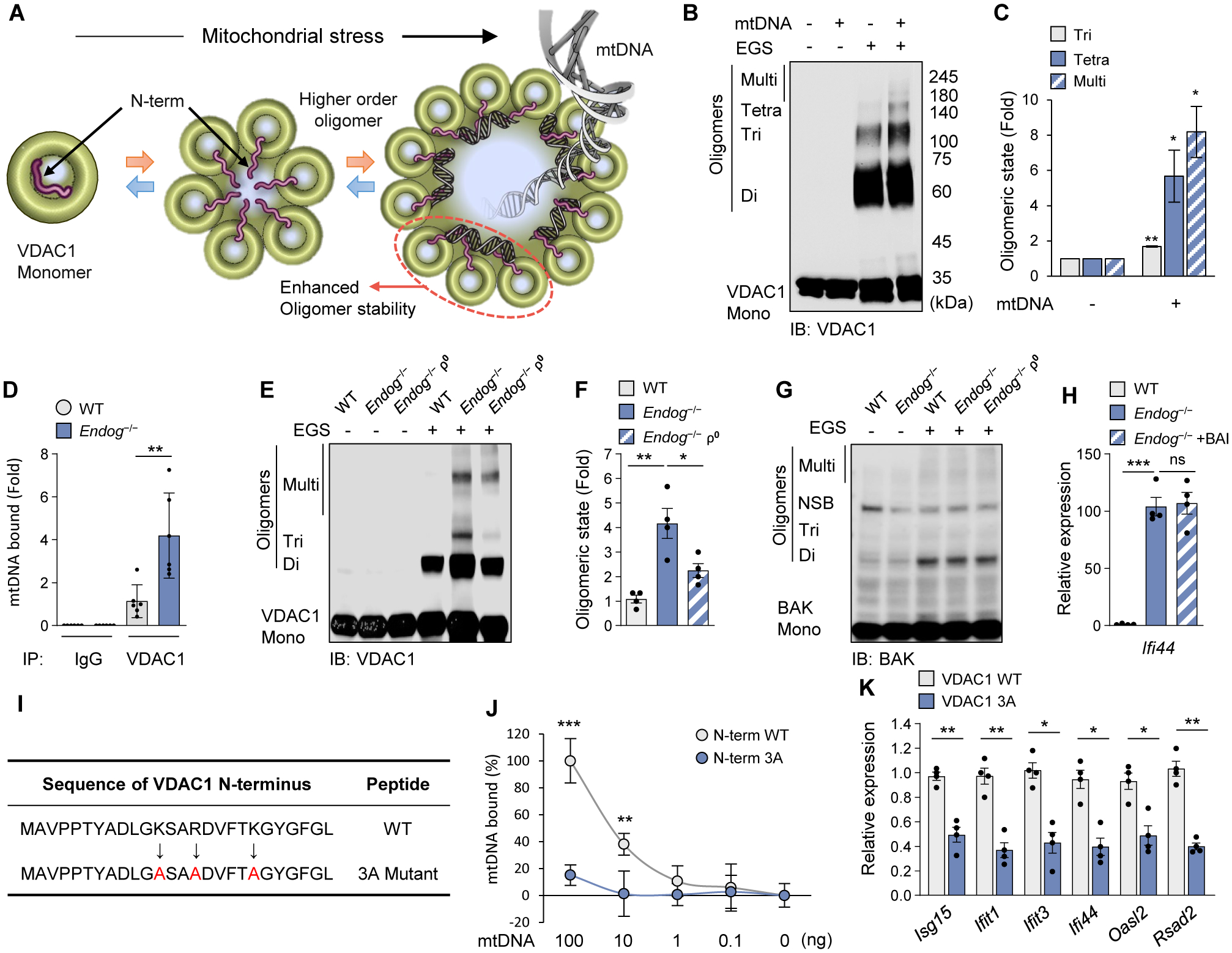
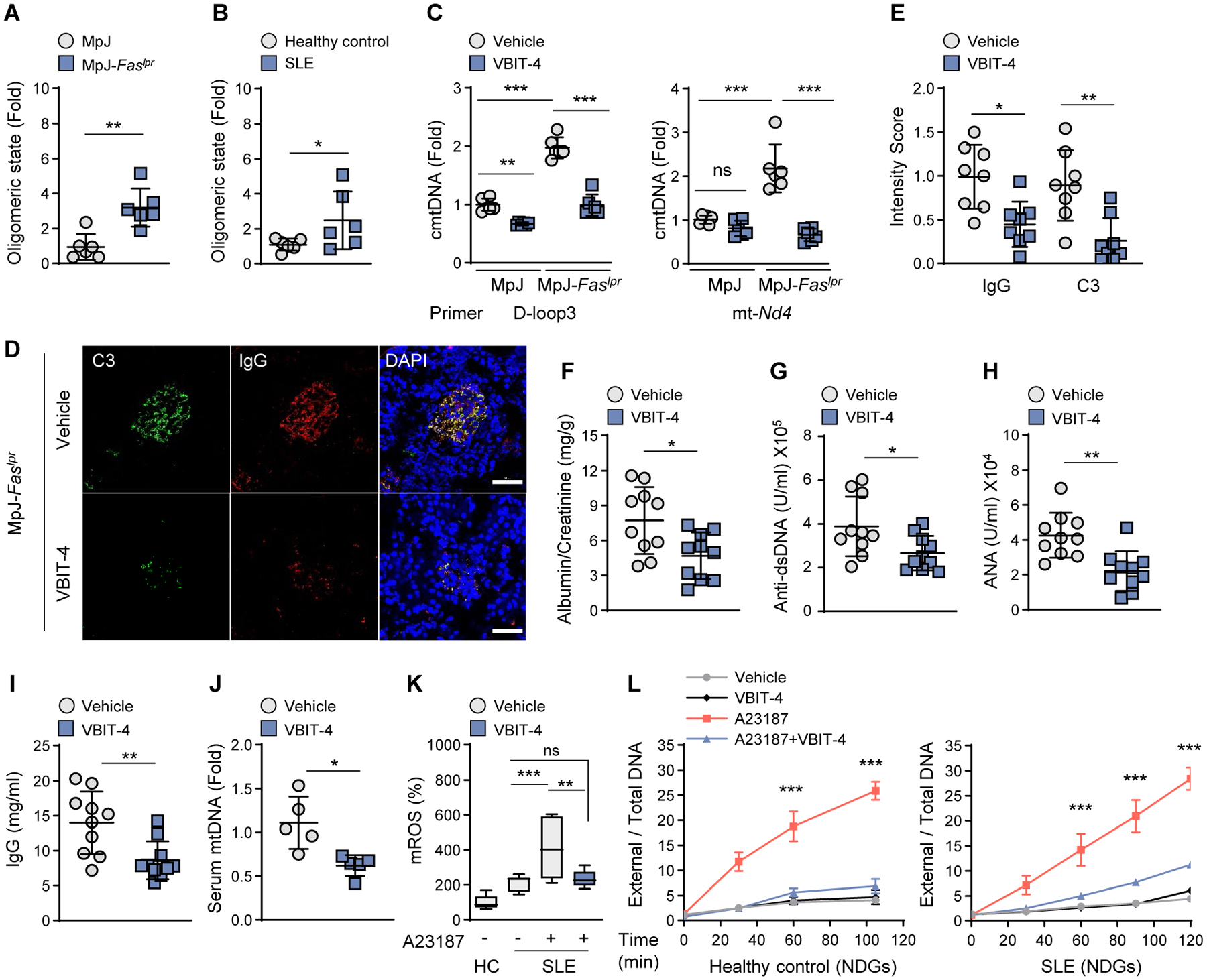
Mitochondrial stress releases mitochondrial DNA (mtDNA) into the cytosol, thereby triggering the type Ι interferon (IFN) response. Mitochondrial outer membrane permeabilization, which is required for mtDNA release, has been extensively studied in apoptotic cells, but little is known about its role in live cells. We found that oxidatively stressed mitochondria release short mtDNA fragments via pores formed by the voltage-dependent anion channel (VDAC) oligomers in the mitochondrial outer membrane. Furthermore, the positively charged residues in the N-terminal domain of VDAC1 interact with mtDNA, promoting VDAC1 oligomerization. The VDAC oligomerization inhibitor VBIT-4 decreases mtDNA release, IFN signaling, neutrophil extracellular traps, and disease severity in a mouse model of systemic lupus erythematosus. Thus, inhibiting VDAC oligomerization is a potential therapeutic approach for diseases associated with mtDNA release.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Mitochondrial DNA promotes autoimmunity.Science. 2019 Dec 20;366(6472):1445-1446. doi: 10.1126/science.aaz9308. Science. 2019. PMID: 31857466 No abstract available.
References
-
- McArthur K et al., BAK/BAX macropores facilitate mitochondrial herniation and mtDNA efflux during apoptosis. Science 359, (2018). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
