

Helix 8 is the essential structural motif of mechanosensitive GPCRs
- PMID: 31857598
- PMCID: PMC6923424
- DOI: 10.1038/s41467-019-13722-0
Helix 8 is the essential structural motif of mechanosensitive GPCRs
Abstract
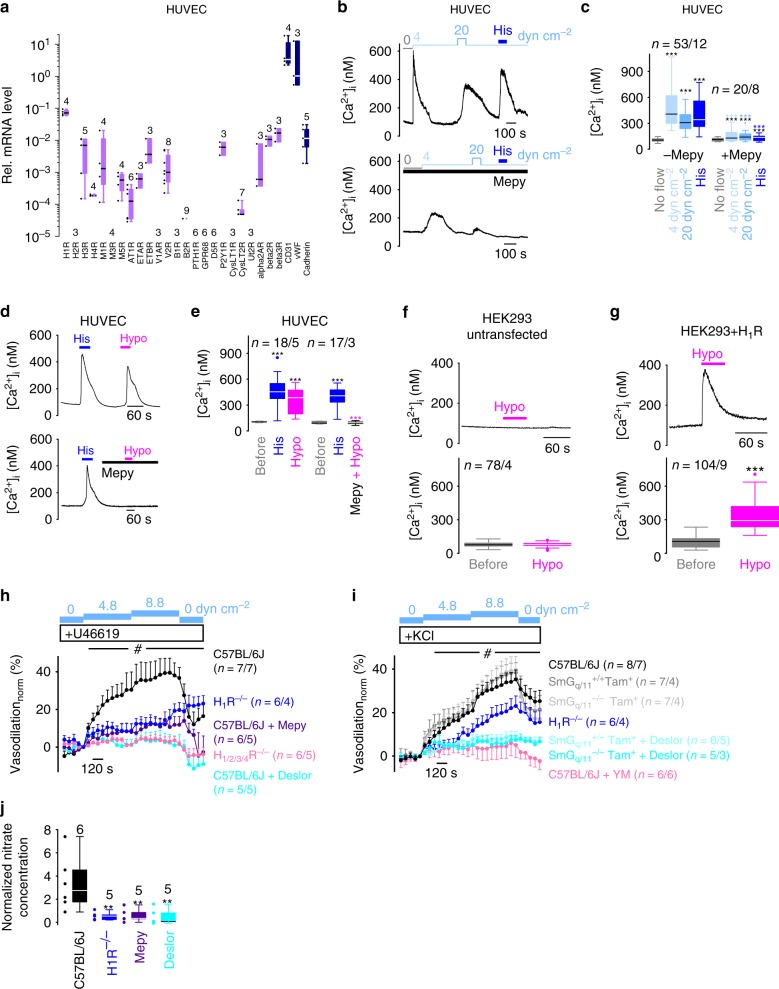
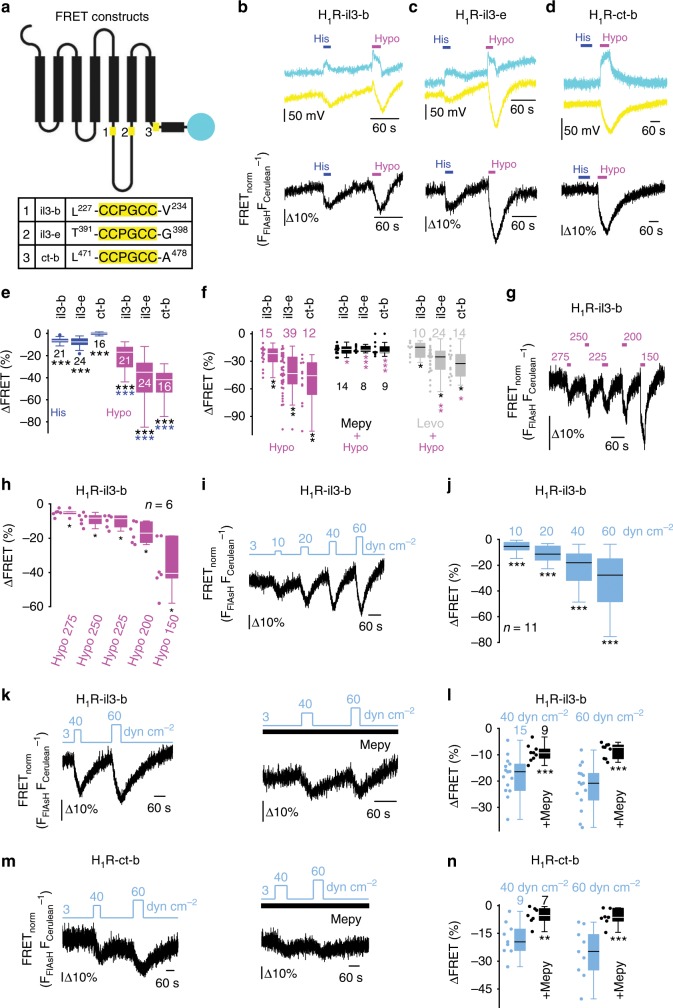
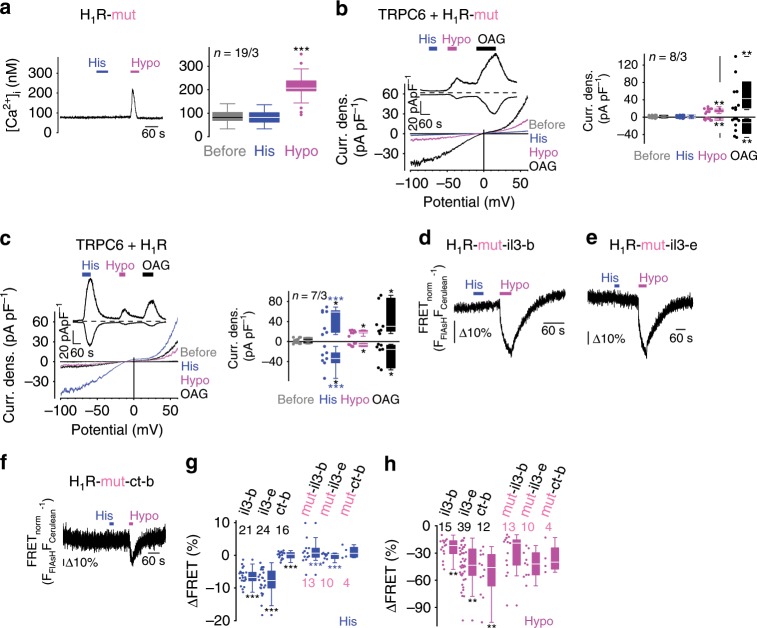
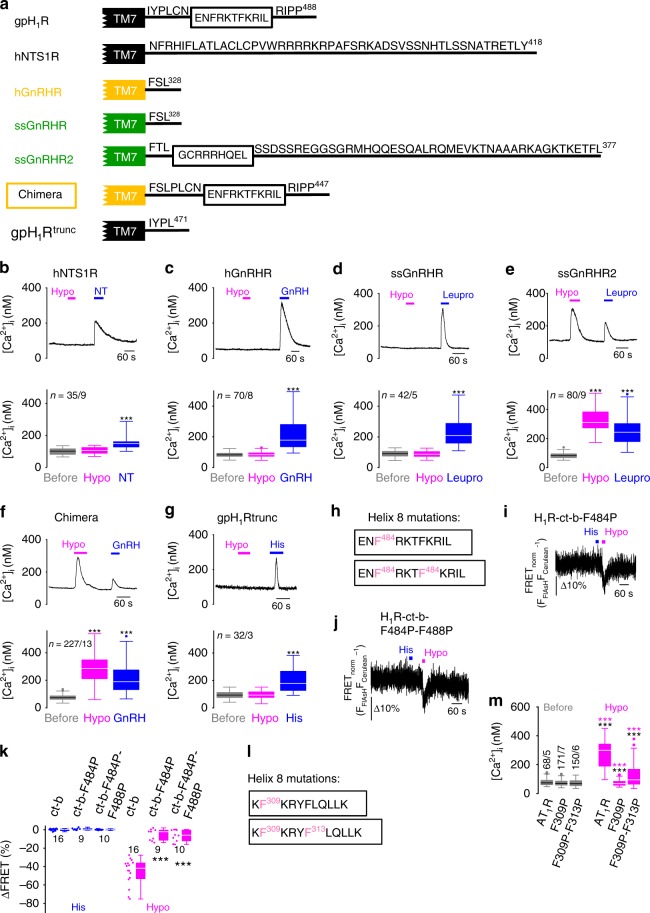
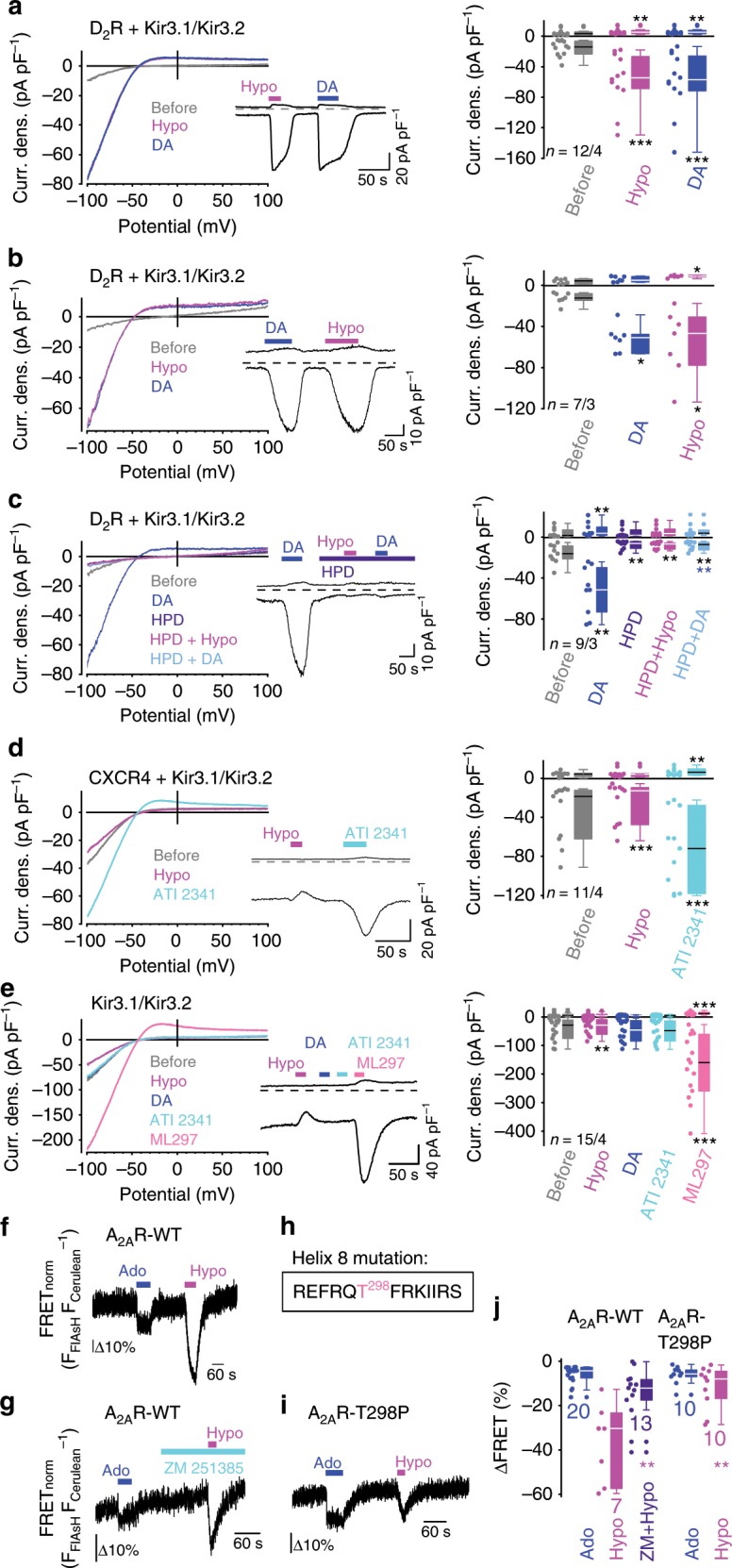
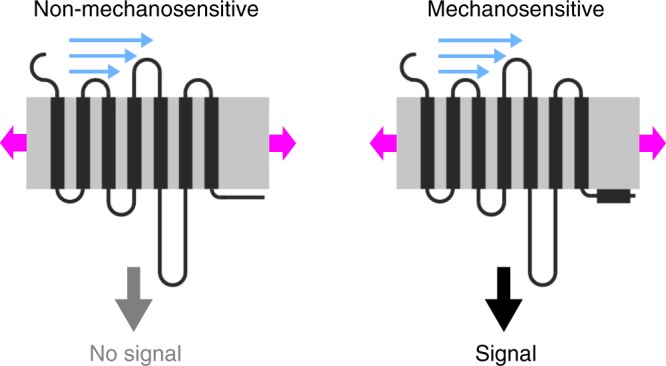
G-protein coupled receptors (GPCRs) are versatile cellular sensors for chemical stimuli, but also serve as mechanosensors involved in various (patho)physiological settings like vascular regulation, cardiac hypertrophy and preeclampsia. However, the molecular mechanisms underlying mechanically induced GPCR activation have remained elusive. Here we show that mechanosensitive histamine H1 receptors (H1Rs) are endothelial sensors of fluid shear stress and contribute to flow-induced vasodilation. At the molecular level, we observe that H1Rs undergo stimulus-specific patterns of conformational changes suggesting that mechanical forces and agonists induce distinct active receptor conformations. GPCRs lacking C-terminal helix 8 (H8) are not mechanosensitive, and transfer of H8 to non-responsive GPCRs confers, while removal of H8 precludes, mechanosensitivity. Moreover, disrupting H8 structural integrity by amino acid exchanges impairs mechanosensitivity. Altogether, H8 is the essential structural motif endowing GPCRs with mechanosensitivity. These findings provide a mechanistic basis for a better understanding of the roles of mechanosensitive GPCRs in (patho)physiology.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Helix 8 - Putting a spring in mechano-sensing.Cell Calcium. 2020 May;87:102192. doi: 10.1016/j.ceca.2020.102192. Epub 2020 Mar 12. Cell Calcium. 2020. PMID: 32200169 No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
