

Evolutionary classification of CRISPR-Cas systems: a burst of class 2 and derived variants
- PMID: 31857715
- PMCID: PMC8905525
- DOI: 10.1038/s41579-019-0299-x
Evolutionary classification of CRISPR-Cas systems: a burst of class 2 and derived variants
Abstract
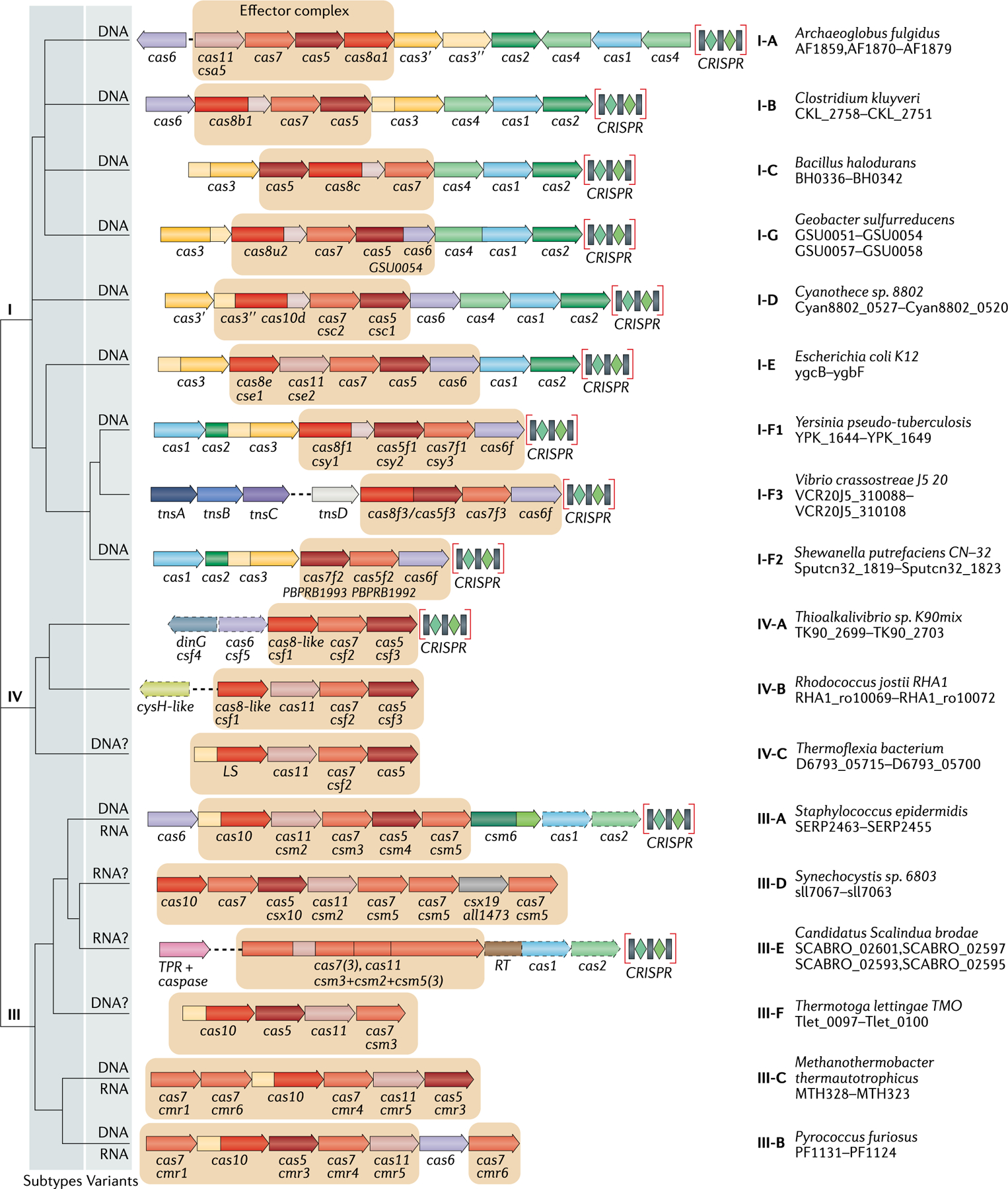
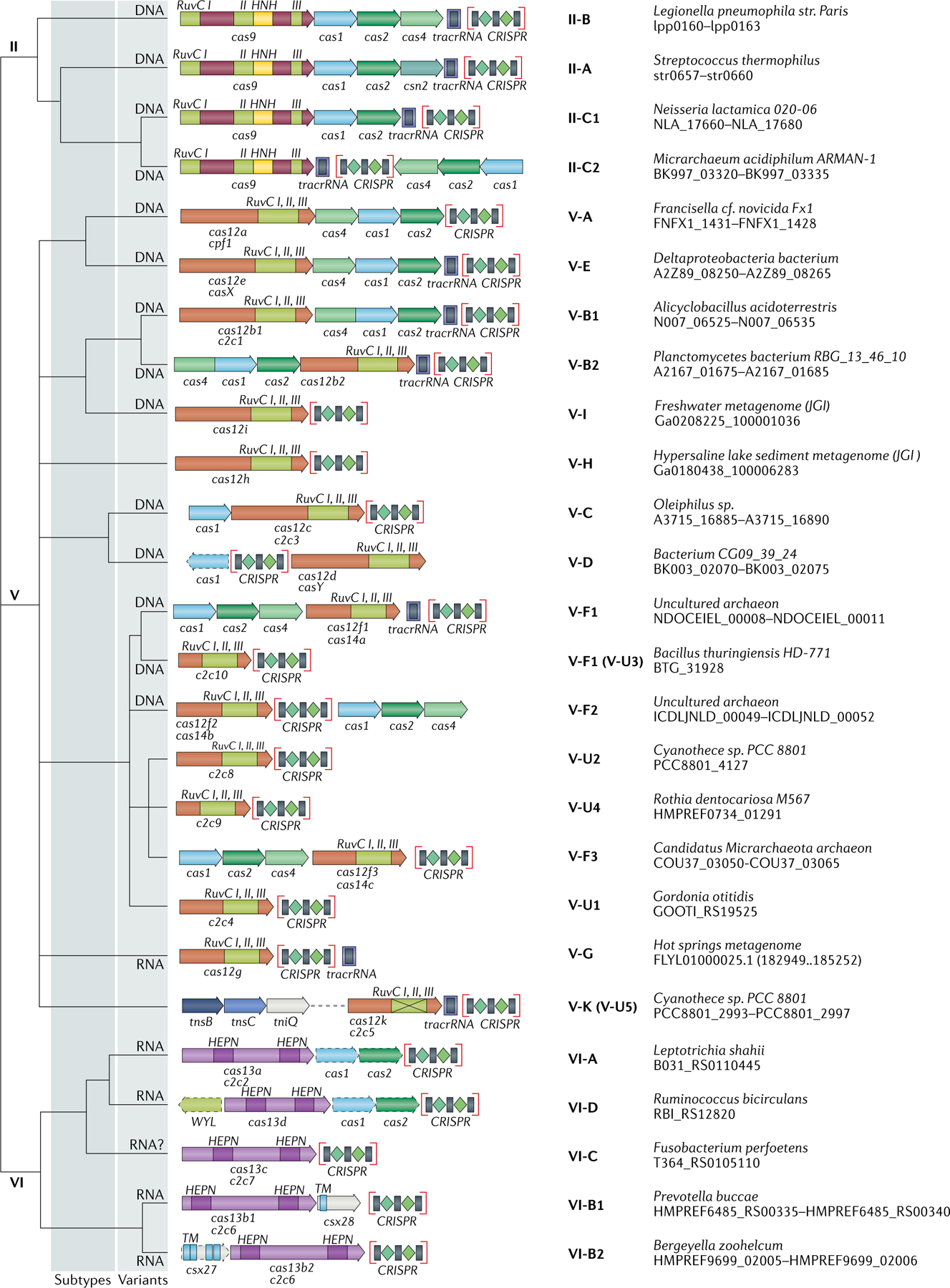
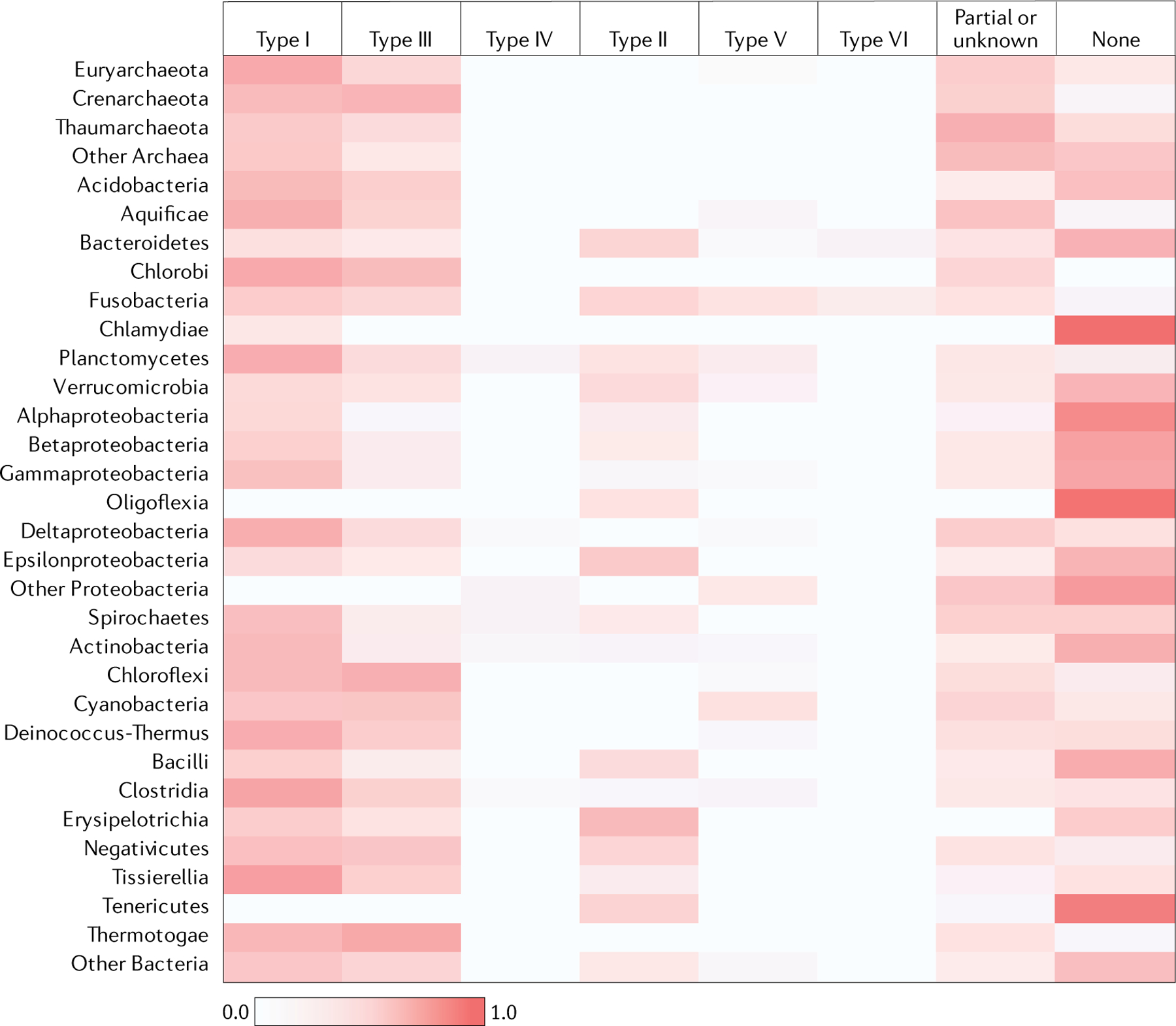
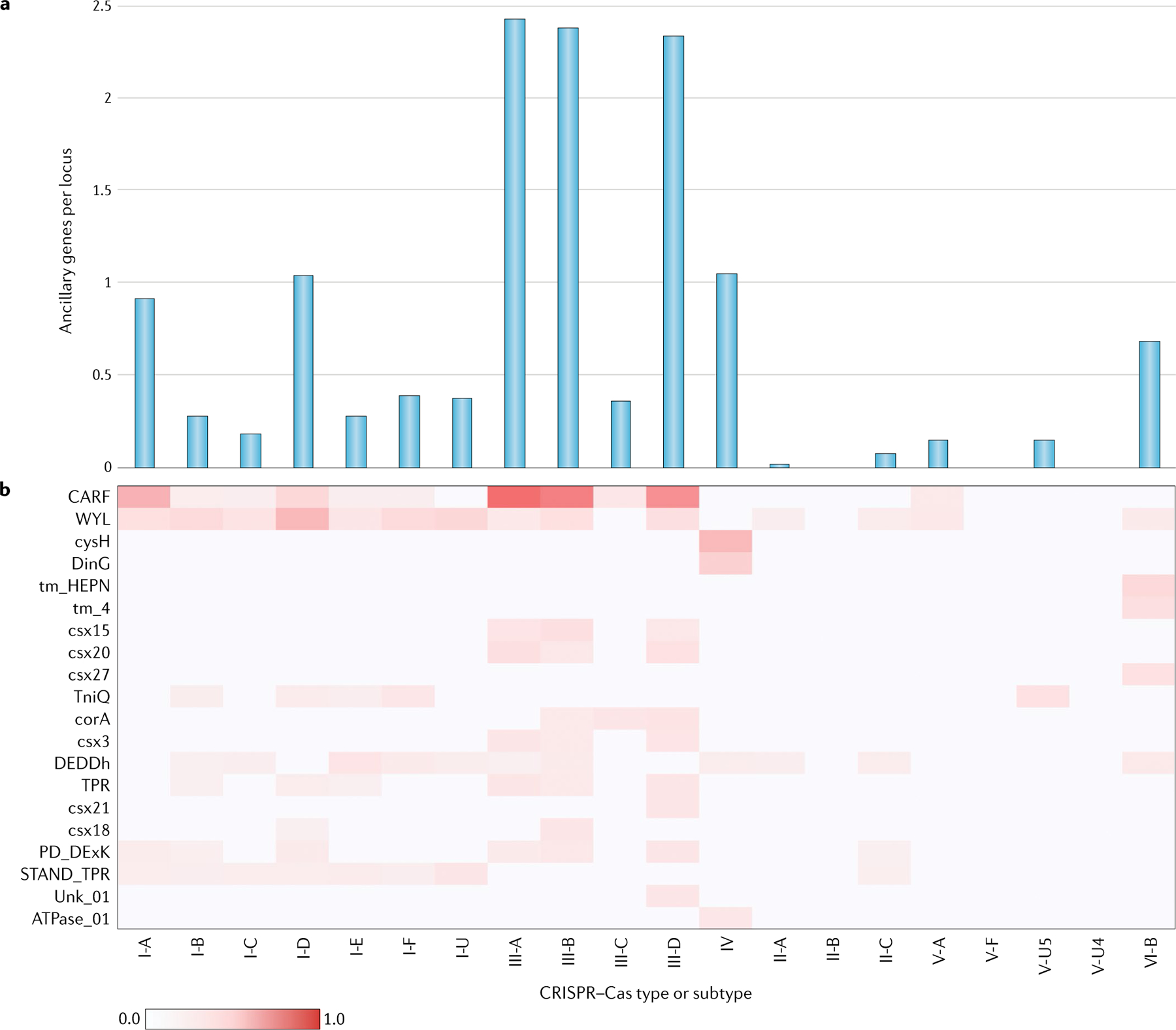
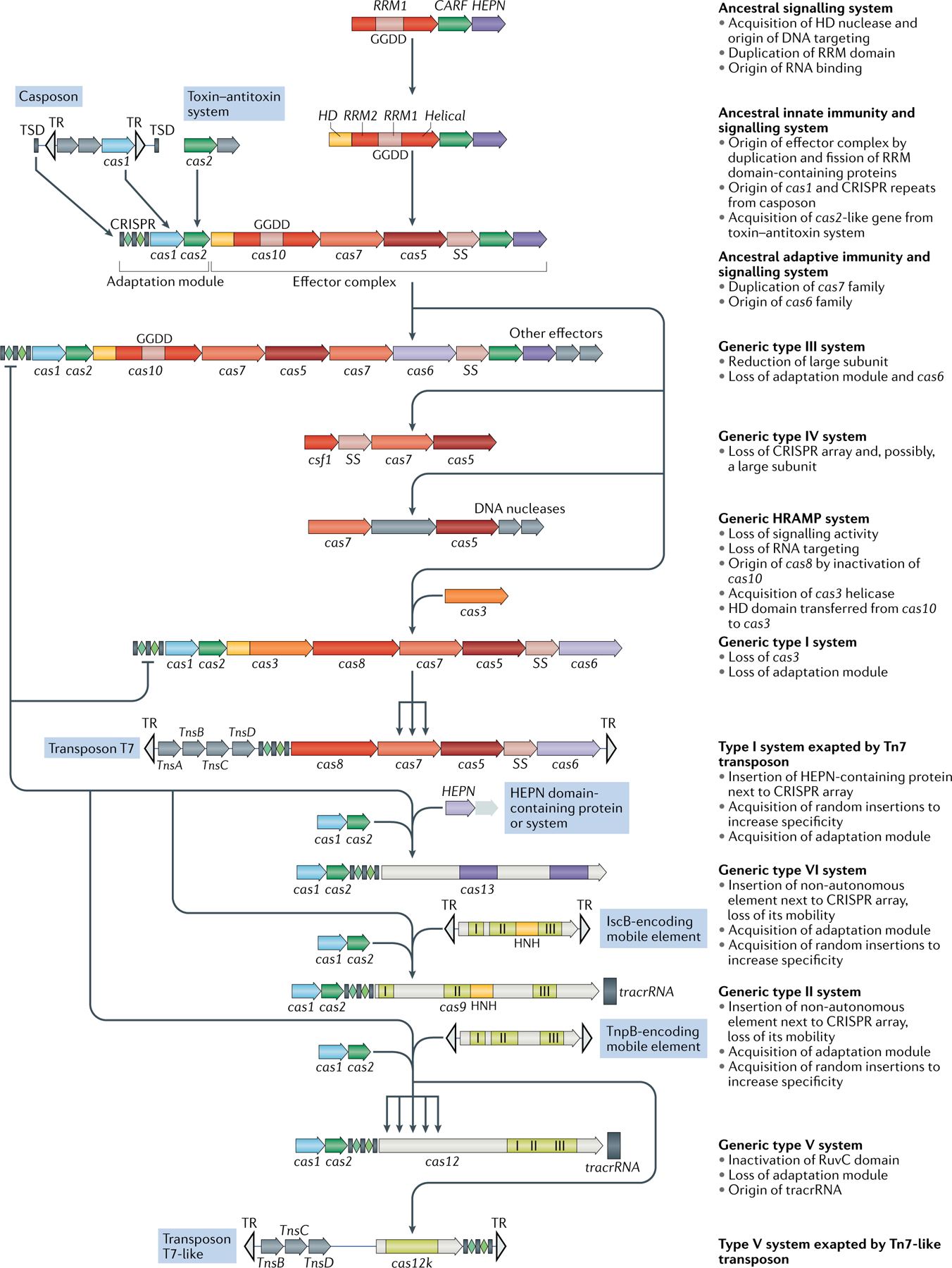
The number and diversity of known CRISPR-Cas systems have substantially increased in recent years. Here, we provide an updated evolutionary classification of CRISPR-Cas systems and cas genes, with an emphasis on the major developments that have occurred since the publication of the latest classification, in 2015. The new classification includes 2 classes, 6 types and 33 subtypes, compared with 5 types and 16 subtypes in 2015. A key development is the ongoing discovery of multiple, novel class 2 CRISPR-Cas systems, which now include 3 types and 17 subtypes. A second major novelty is the discovery of numerous derived CRISPR-Cas variants, often associated with mobile genetic elements that lack the nucleases required for interference. Some of these variants are involved in RNA-guided transposition, whereas others are predicted to perform functions distinct from adaptive immunity that remain to be characterized experimentally. The third highlight is the discovery of numerous families of ancillary CRISPR-linked genes, often implicated in signal transduction. Together, these findings substantially clarify the functional diversity and evolutionary history of CRISPR-Cas.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
-
- Mohanraju P et al. Diverse evolutionary roots and mechanistic variations of the CRISPR–Cas systems. Science 353, aad5147 (2016). - PubMed
-
- Jackson SA et al. CRISPR–Cas: adapting to change. Science 356, eaal5056 (2017). - PubMed
-
- Barrangou R & Horvath P A decade of discovery: CRISPR functions and applications. Nat. Microbiol 2, 17092 (2017). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
