

Restricted and non-essential redundancy of RNAi and piRNA pathways in mouse oocytes
- PMID: 31860668
- PMCID: PMC6944382
- DOI: 10.1371/journal.pgen.1008261
Restricted and non-essential redundancy of RNAi and piRNA pathways in mouse oocytes
Abstract
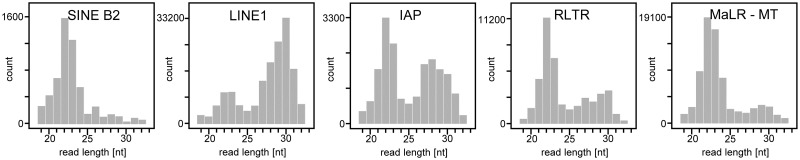
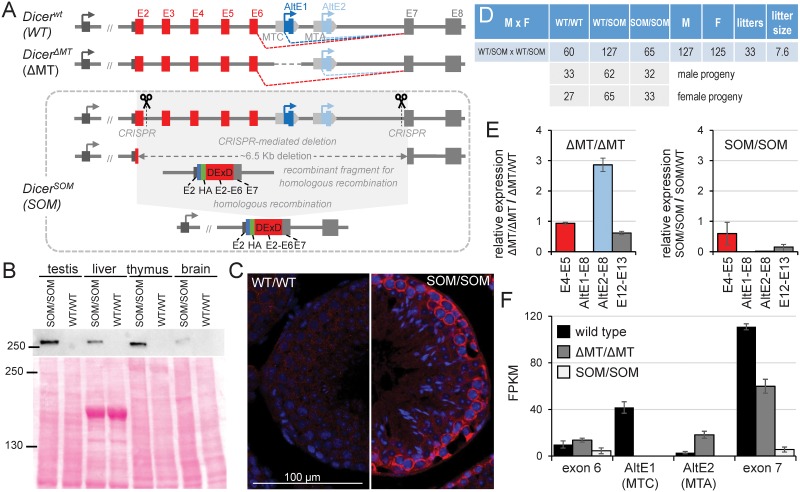
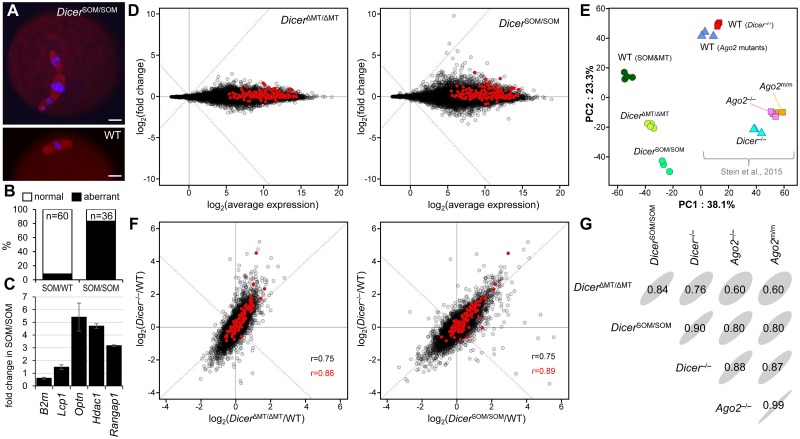
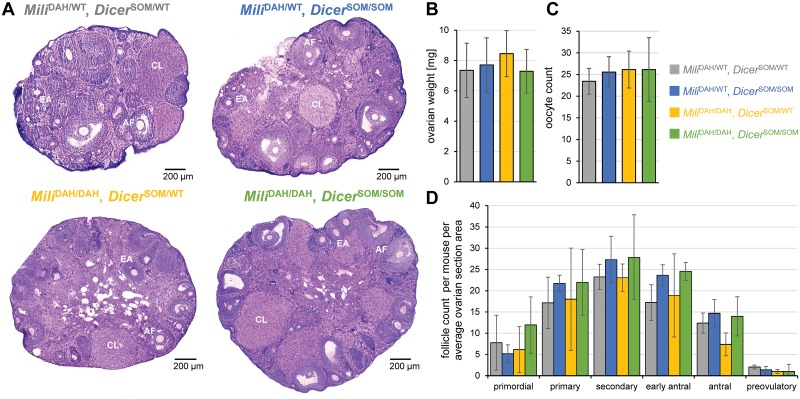
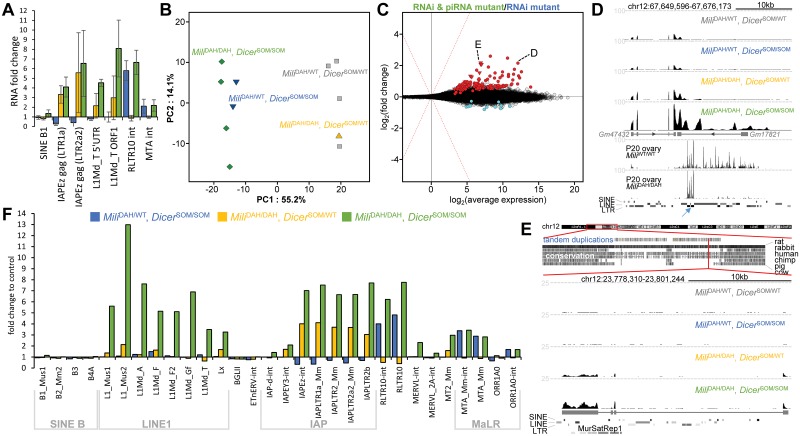
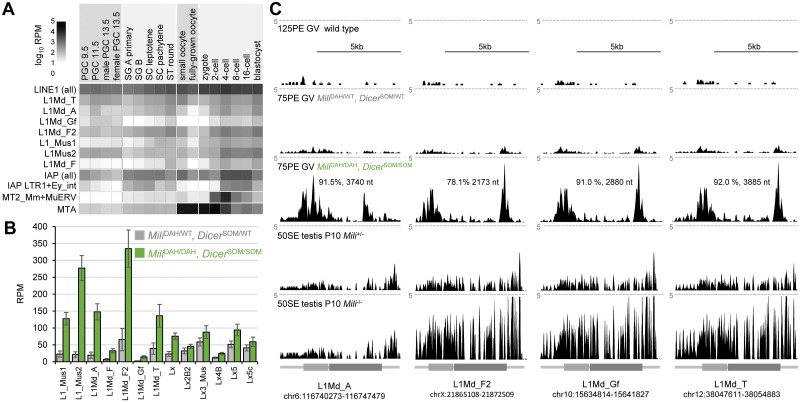
Germline genome defense evolves to recognize and suppress retrotransposons. One of defensive mechanisms is the PIWI-associated RNA (piRNA) pathway, which employs small RNAs for sequence-specific repression. The loss of the piRNA pathway in mice causes male sterility while females remain fertile. Unlike spermatogenic cells, mouse oocytes posses also RNA interference (RNAi), another small RNA pathway capable of retrotransposon suppression. To examine whether RNAi compensates the loss of the piRNA pathway, we produced a new RNAi pathway mutant DicerSOM and crossed it with a catalytically-dead mutant of Mili, an essential piRNA gene. Normal follicular and oocyte development in double mutants showed that RNAi does not suppress a strong ovarian piRNA knock-out phenotype. However, we observed redundant and non-redundant targeting of specific retrotransposon families illustrating stochasticity of recognition and targeting of invading retrotransposons. Intracisternal A Particle retrotransposon was mainly targeted by the piRNA pathway, MaLR and RLTR10 retrotransposons were targeted mainly by RNAi. Double mutants showed accumulations of LINE-1 retrotransposon transcripts. However, we did not find strong evidence for transcriptional activation and mobilization of retrotransposition competent LINE-1 elements suggesting that while both defense pathways are simultaneously expendable for ovarian oocyte development, yet another transcriptional silencing mechanism prevents mobilization of LINE-1 elements.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Craig NL, Chandler M, Gellert M, Lambowitz AM, Rice PA, Sandmeyer SB. Mobile DNA III. Craig NL, editor: AMS press; 2015.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
