

Targeting proline in (phospho)proteomics
- PMID: 31863553
- PMCID: PMC7496877
- DOI: 10.1111/febs.15190
Targeting proline in (phospho)proteomics
Abstract
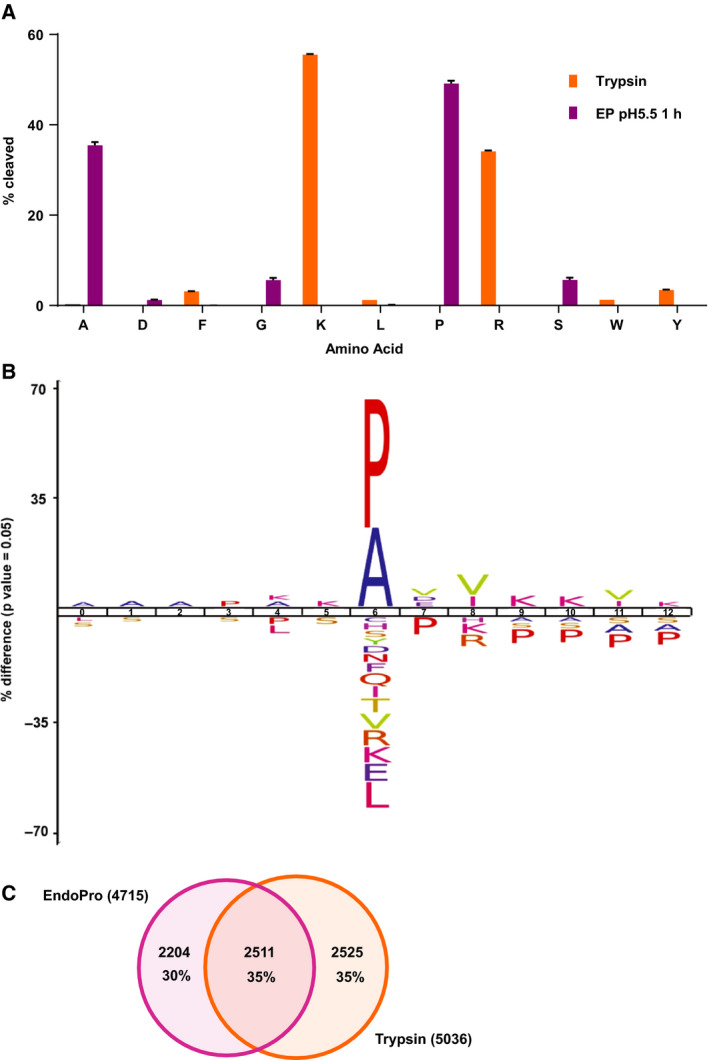
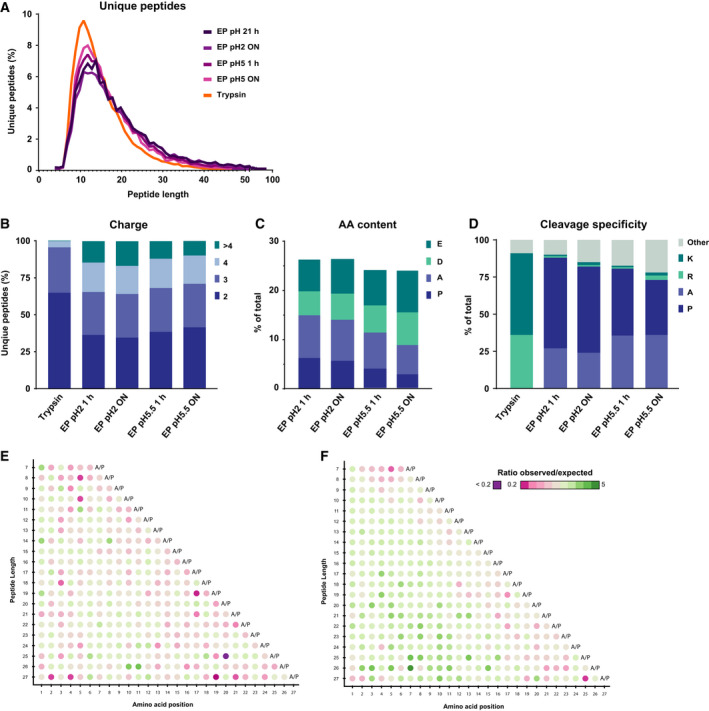
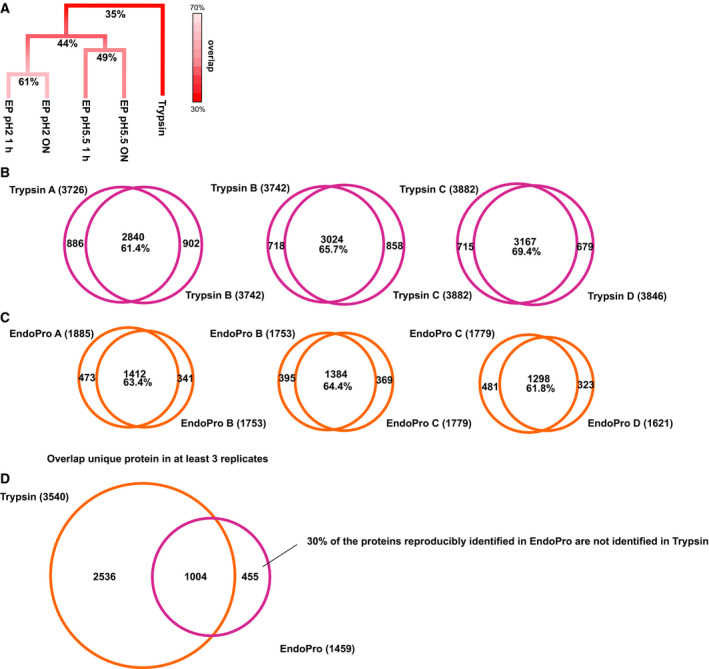
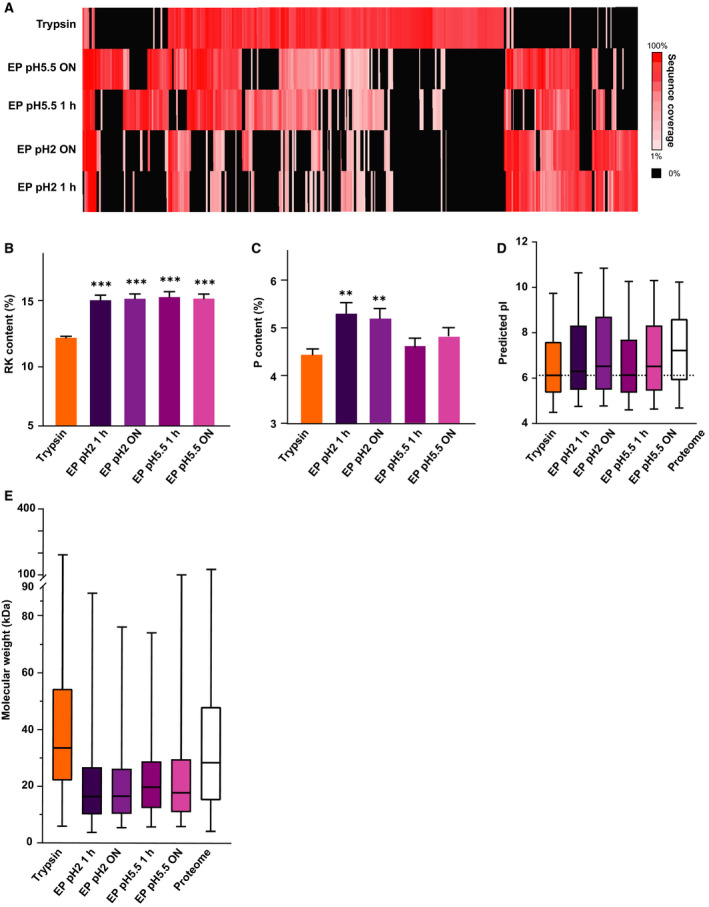
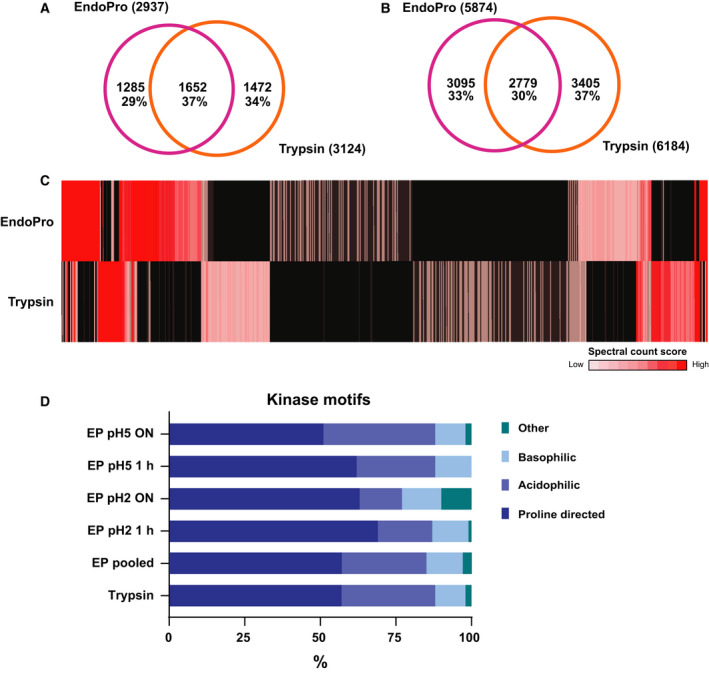
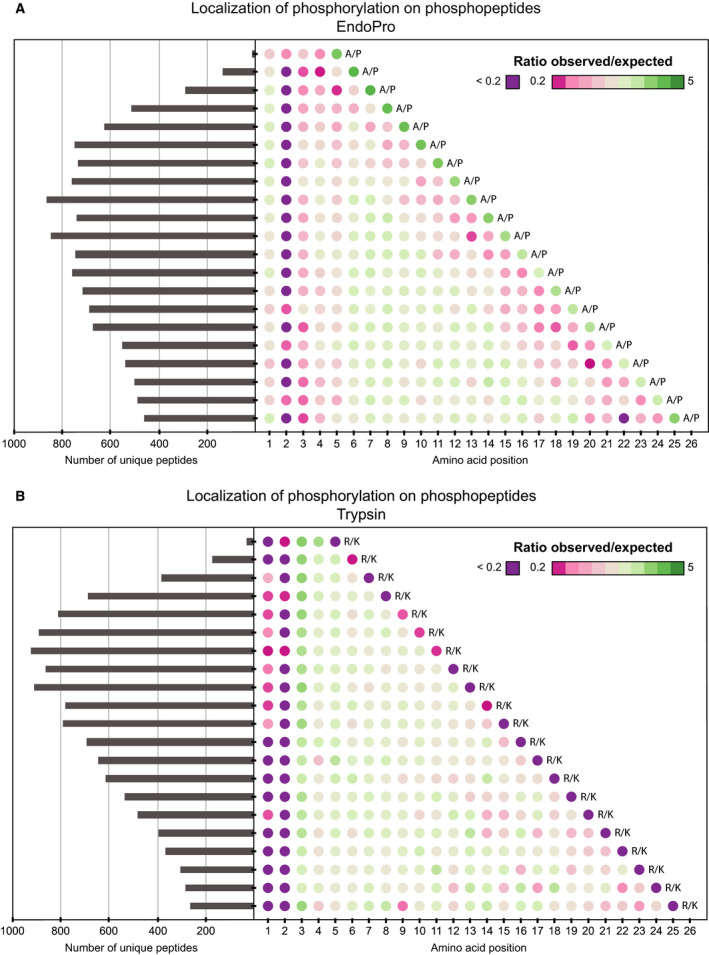
Mass spectrometry-based proteomics experiments typically start with the digestion of proteins using trypsin, chosen because of its high specificity, availability, and ease of use. It has become apparent that the sole use of trypsin may impose certain limits on our ability to grasp the full proteome, missing out particular sites of post-translational modifications, protein segments, or even subsets of proteins. To tackle this problem, alternative proteases have been introduced and shown to lead to an increase in the detectable (phospho)proteome. Here, we argue that there may be further room for improvement and explore the protease EndoPro. For optimal peptide identification rates, we explored multiple peptide fragmentation techniques (HCD, ETD, and EThcD) and employed Byonic as search algorithm. We obtain peptide IDs for about 40% of the MS2 spectra (66% for trypsin). EndoPro cleaves with high specificity at the C-terminal site of Pro and Ala residues and displays activity in a broad pH range, where we focused on its performance at pH = 2 and 5.5. The proteome coverage of EndoPro at these two pH values is rather distinct, and also complementary to the coverage obtained with trypsin. As about 40% of mammalian protein phosphorylations are proline-directed, we also explored the performance of EndoPro in phosphoproteomics. EndoPro extends the coverable phosphoproteome substantially, whereby both the, at pH = 2 and 5.5, acquired phosphoproteomes are complementary to each other and to the phosphoproteome obtained using trypsin. Hence, EndoPro is a powerful tool to exploit in (phospho)proteomics applications.
Keywords: (phospho)proteomics; EndoPro; mass spectrometry; proline effect; protease.
© 2019 The Authors. The FEBS Journal published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Conflict of interest statement
The authors declare the following competing financial interest(s): M.A. and M.O. are DSM employees. DSM sells An‐PEP for food applications. MB is the founder and employee of ProteinMetrics. ProteinMetrics develops and commercializes the Byonic software.
Figures
Similar articles
-
ProAlanase is an Effective Alternative to Trypsin for Proteomics Applications and Disulfide Bond Mapping.Mol Cell Proteomics. 2020 Dec;19(12):2139-2157. doi: 10.1074/mcp.TIR120.002129. Epub 2020 Oct 5. Mol Cell Proteomics. 2020. PMID: 33020190 Free PMC article.
-
Six alternative proteases for mass spectrometry-based proteomics beyond trypsin.Nat Protoc. 2016 May;11(5):993-1006. doi: 10.1038/nprot.2016.057. Epub 2016 Apr 28. Nat Protoc. 2016. PMID: 27123950
-
Proteomics beyond trypsin.FEBS J. 2015 Jul;282(14):2612-26. doi: 10.1111/febs.13287. Epub 2015 Apr 14. FEBS J. 2015. PMID: 25823410 Review.
-
Opposite Electron-Transfer Dissociation and Higher-Energy Collisional Dissociation Fragmentation Characteristics of Proteolytic K/R(X)n and (X)nK/R Peptides Provide Benefits for Peptide Sequencing in Proteomics and Phosphoproteomics.J Proteome Res. 2017 Feb 3;16(2):852-861. doi: 10.1021/acs.jproteome.6b00825. Epub 2016 Dec 2. J Proteome Res. 2017. PMID: 28111955
-
Getting intimate with trypsin, the leading protease in proteomics.Mass Spectrom Rev. 2013 Nov-Dec;32(6):453-65. doi: 10.1002/mas.21376. Epub 2013 Jun 15. Mass Spectrom Rev. 2013. PMID: 23775586 Review.
Cited by
-
Native Structural and Functional Proteoform Characterization of the Prolyl-Alanyl-Specific Endoprotease EndoPro from Aspergillus niger.J Proteome Res. 2021 Oct 1;20(10):4875-4885. doi: 10.1021/acs.jproteome.1c00663. Epub 2021 Sep 13. J Proteome Res. 2021. PMID: 34515489 Free PMC article. Review.
-
Structural and time-resolved mechanistic investigations of protein hydrolysis by the acidic proline-specific endoprotease from Aspergillus niger.Protein Sci. 2024 Jan;33(1):e4856. doi: 10.1002/pro.4856. Protein Sci. 2024. PMID: 38059672 Free PMC article.
-
The lysosomal endopeptidases Cathepsin D and L are selective and effective proteases for the middle-down characterization of antibodies.FEBS J. 2021 Sep;288(18):5389-5405. doi: 10.1111/febs.15813. Epub 2021 Mar 27. FEBS J. 2021. PMID: 33713388 Free PMC article.
-
Phosphoproteomics: a valuable tool for uncovering molecular signaling in cancer cells.Expert Rev Proteomics. 2021 Aug;18(8):661-674. doi: 10.1080/14789450.2021.1976152. Epub 2021 Sep 16. Expert Rev Proteomics. 2021. PMID: 34468274 Free PMC article. Review.
-
Postproline Cleaving Enzymes also Show Specificity to Reduced Cysteine.Anal Chem. 2024 Dec 3;96(48):19084-19092. doi: 10.1021/acs.analchem.4c04277. Epub 2024 Nov 19. Anal Chem. 2024. PMID: 39560312 Free PMC article.
References
-
- Cohen P (2001) The role of protein phosphorylation in human health and disease. Eur J Biochem 268, 5001–5010. - PubMed
-
- Hanash S (2003) Disease proteomics. Nature 422, 226–232. - PubMed
-
- Mann M, Kulak NA, Nagaraj N & Cox J (2013) The coming age of complete, accurate, and ubiquitous proteomes. Mol Cell 49, 583–590. - PubMed
-
- Steen H & Mann M (2004) The abc's (and xyz's) of peptide sequencing. Nat Rev Mol Cell Biol 5, 699–711. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
