

The microbiota influences the Drosophila melanogaster life history strategy
- PMID: 31863671
- PMCID: PMC7507235
- DOI: 10.1111/mec.15344
The microbiota influences the Drosophila melanogaster life history strategy
Abstract
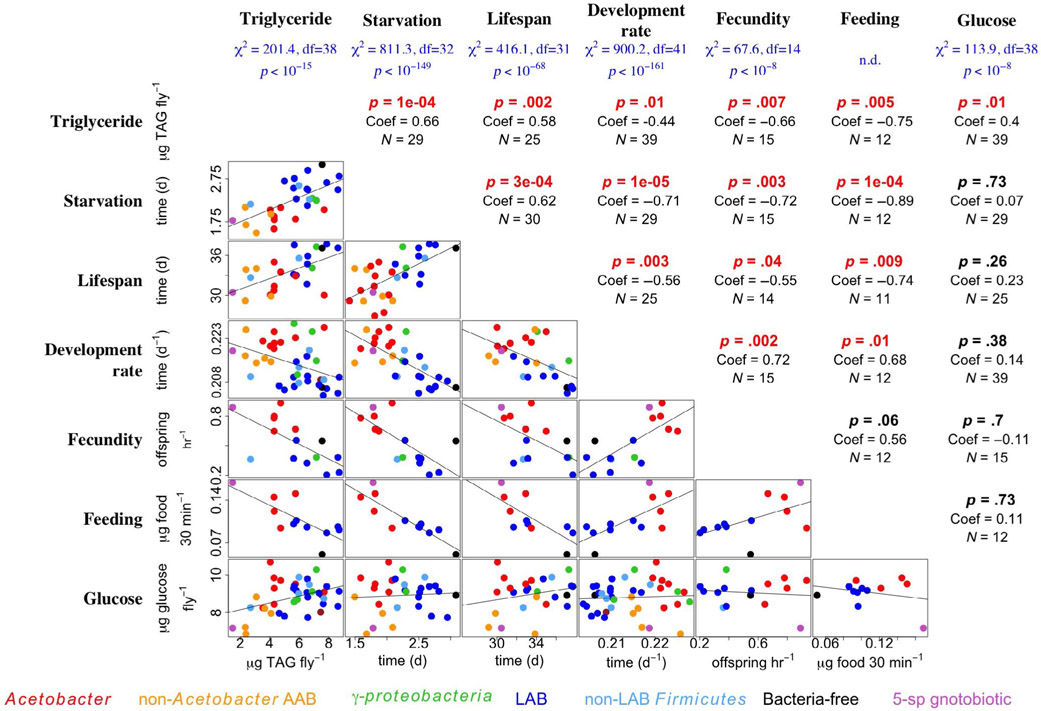
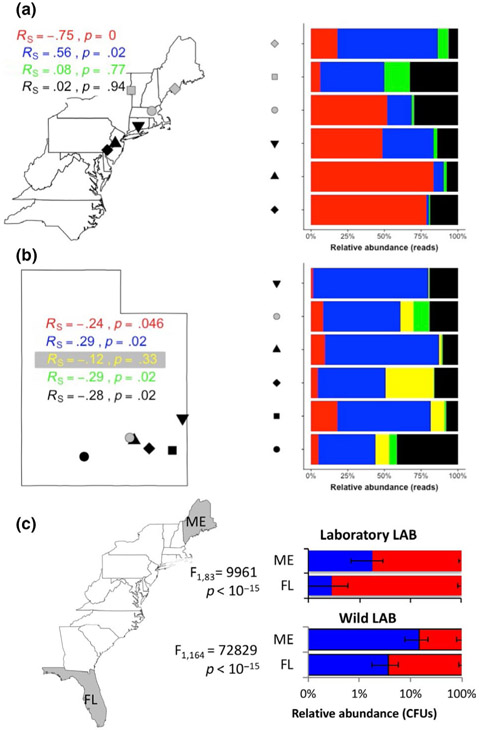
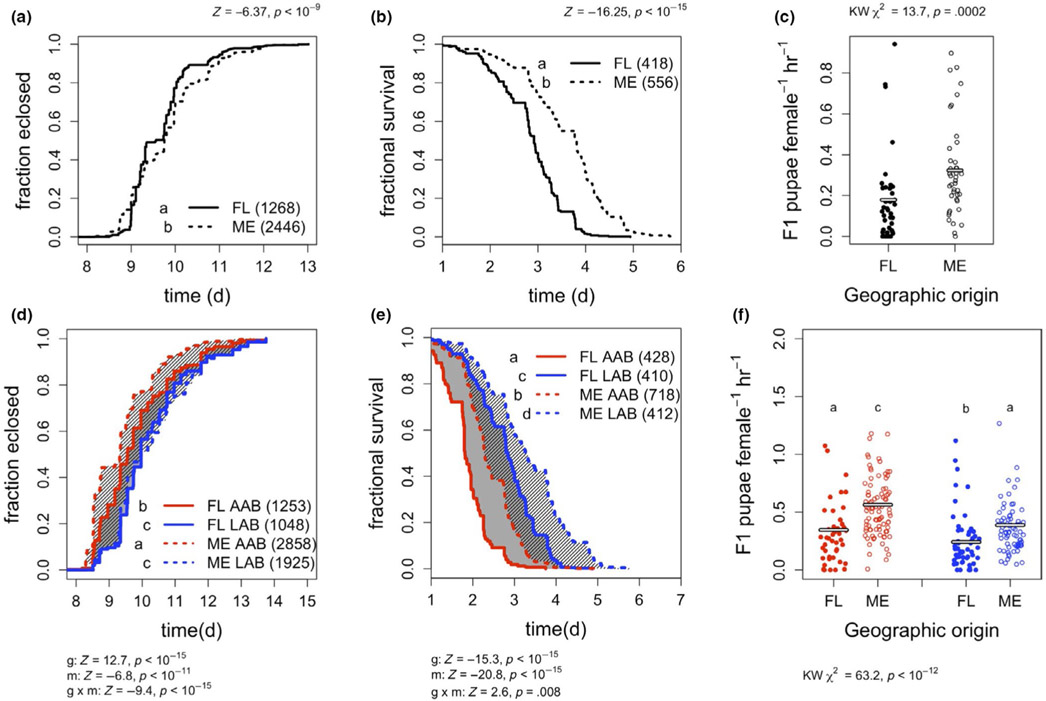
Organisms are locally adapted when members of a population have a fitness advantage in one location relative to conspecifics in other geographies. For example, across latitudinal gradients, some organisms may trade off between traits that maximize fitness components in one, but not both, of somatic maintenance or reproductive output. Latitudinal gradients in life history strategies are traditionally attributed to environmental selection on an animal's genotype, without any consideration of the possible impact of associated microorganisms ("microbiota") on life history traits. Here, we show in Drosophila melanogaster, a key model for studying local adaptation and life history strategy, that excluding the microbiota from definitions of local adaptation is a major shortfall. First, we reveal that an isogenic fly line reared with different bacteria varies the investment in early reproduction versus somatic maintenance. Next, we show that in wild fruit flies, the abundance of these same bacteria was correlated with the latitude and life history strategy of the flies, suggesting geographic specificity of the microbiota composition. Variation in microbiota composition of locally adapted D. melanogaster could be attributed to both the wild environment and host genetic selection. Finally, by eliminating or manipulating the microbiota of fly lines collected across a latitudinal gradient, we reveal that host genotype contributes to latitude-specific life history traits independent of the microbiota and that variation in the microbiota can suppress or reverse the differences between locally adapted fly lines. Together, these findings establish the microbiota composition of a model animal as an essential consideration in local adaptation.
Keywords: Drosophila; acetic acid bacteria; lactic acid bacteria; latitude; life history; local adaptation; microbiome; microbiota.
© 2019 John Wiley & Sons Ltd.
Figures
References
-
- Bates D, Mächler M, Bolker B, & Walker S (2015). Fitting linear mixed-effects models using lme4. Journal of Statistical Software, 67(1), 1–48. 10.18637/jss.v067.i01 - DOI
-
- Benson AK, Kelly SA, Legge R, Ma F, Low SJ, Kim J, … Pomp D (2010). Individuality in gut microbiota composition is a complex polygenic trait shaped by multiple environmental and host genetic factors. Proceedings of the National Academy of Sciences of the United States of America, 107(44), 18933–18938. 10.1073/pnas.1007028107 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
