

Bridging Single Neuron Dynamics to Global Brain States
- PMID: 31866837
- PMCID: PMC6908479
- DOI: 10.3389/fnsys.2019.00075
Bridging Single Neuron Dynamics to Global Brain States
Abstract
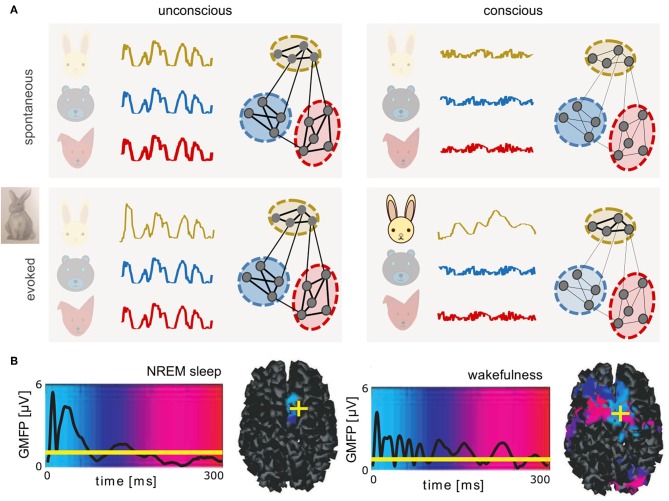
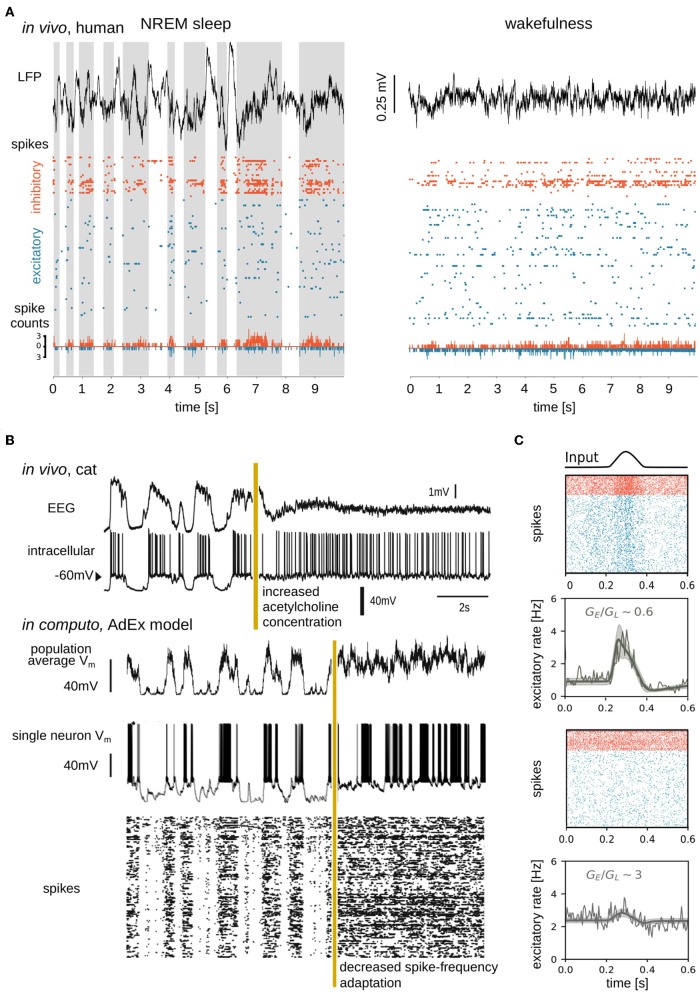
Biological neural networks produce information backgrounds of multi-scale spontaneous activity that become more complex in brain states displaying higher capacities for cognition, for instance, attentive awake versus asleep or anesthetized states. Here, we review brain state-dependent mechanisms spanning ion channel currents (microscale) to the dynamics of brain-wide, distributed, transient functional assemblies (macroscale). Not unlike how microscopic interactions between molecules underlie structures formed in macroscopic states of matter, using statistical physics, the dynamics of microscopic neural phenomena can be linked to macroscopic brain dynamics through mesoscopic scales. Beyond spontaneous dynamics, it is observed that stimuli evoke collapses of complexity, most remarkable over high dimensional, asynchronous, irregular background dynamics during consciousness. In contrast, complexity may not be further collapsed beyond synchrony and regularity characteristic of unconscious spontaneous activity. We propose that increased dimensionality of spontaneous dynamics during conscious states supports responsiveness, enhancing neural networks' emergent capacity to robustly encode information over multiple scales.
Keywords: cerebral cortex; computational neuroscience; coupling; desynchronized; low-dimensional manifold; mean-field models; membrane biophysics; neural network models.
Copyright © 2019 Goldman, Tort-Colet, di Volo, Susin, Bouté, Dali, Carlu, Nghiem, Górski and Destexhe.
Figures
References
LinkOut - more resources
Full Text Sources