

Combined single-cell and spatial transcriptomics reveal the molecular, cellular and spatial bone marrow niche organization
- PMID: 31871321
- PMCID: PMC7610809
- DOI: 10.1038/s41556-019-0439-6
Combined single-cell and spatial transcriptomics reveal the molecular, cellular and spatial bone marrow niche organization
Abstract
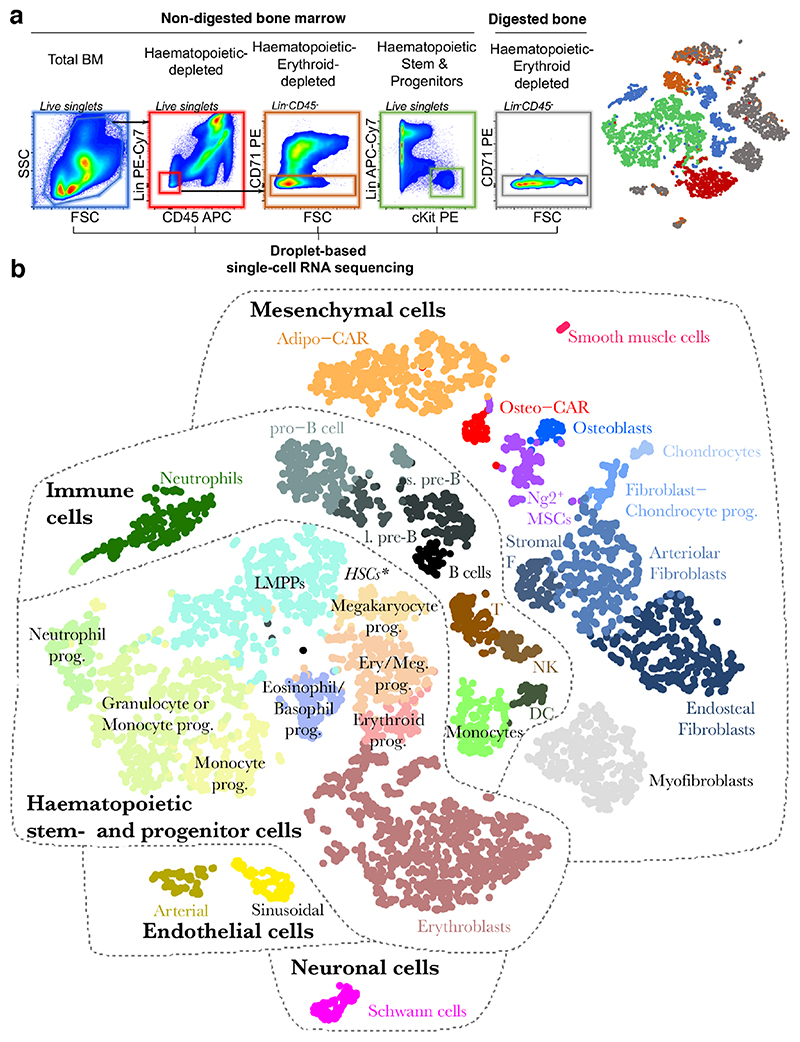
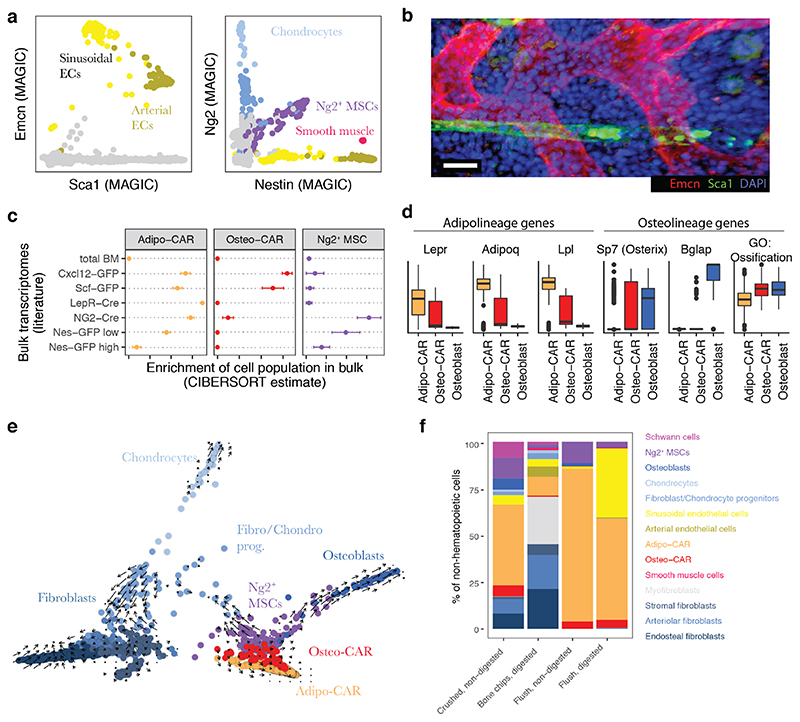
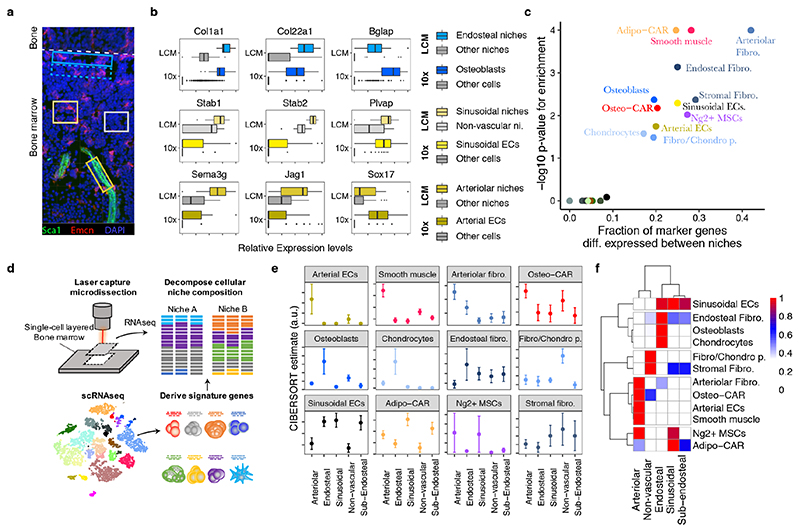
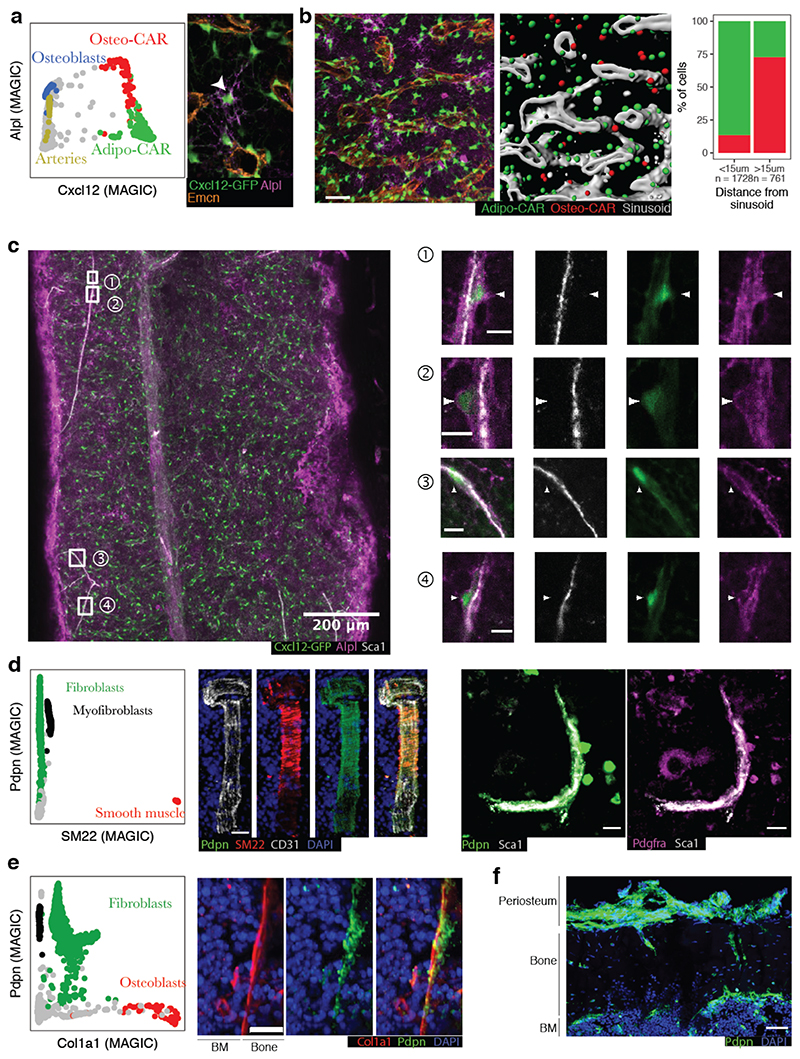
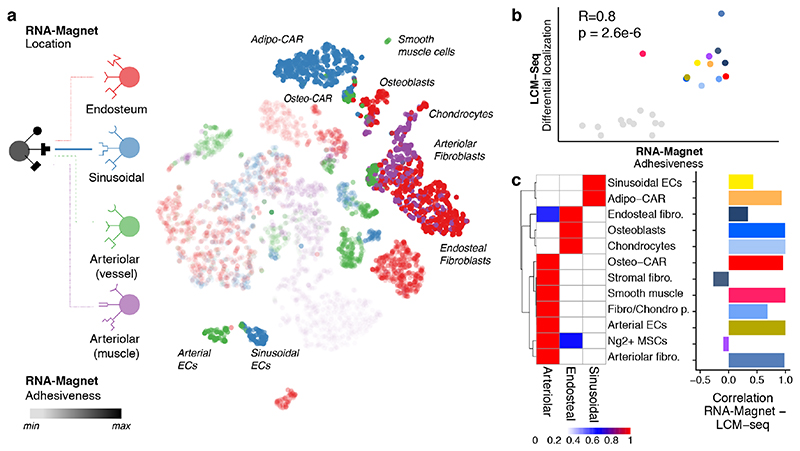
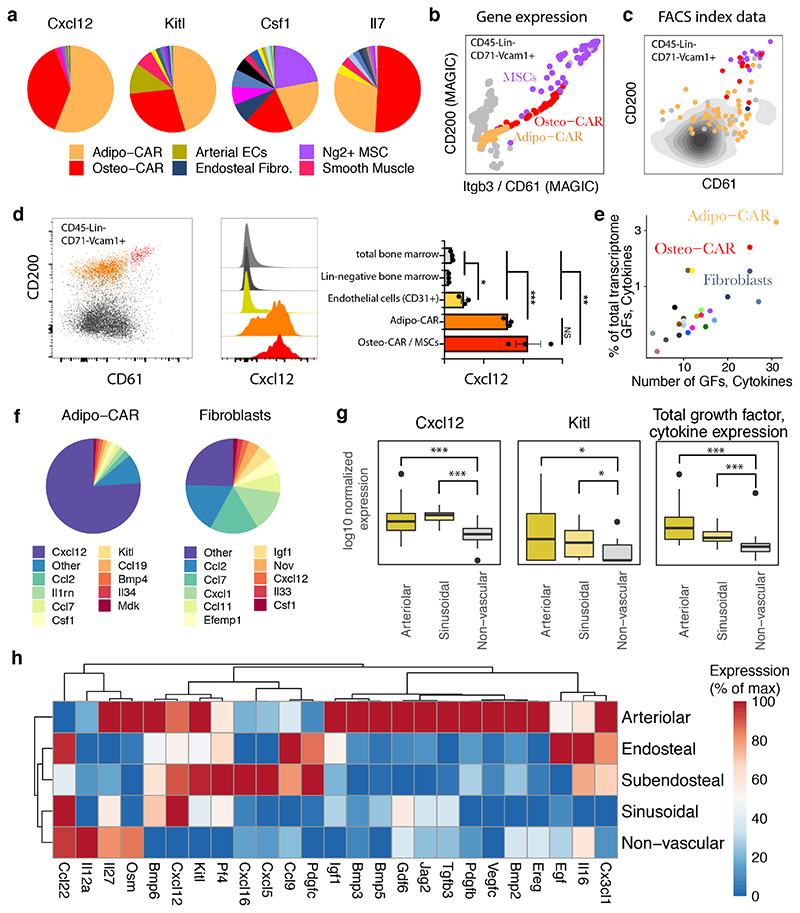
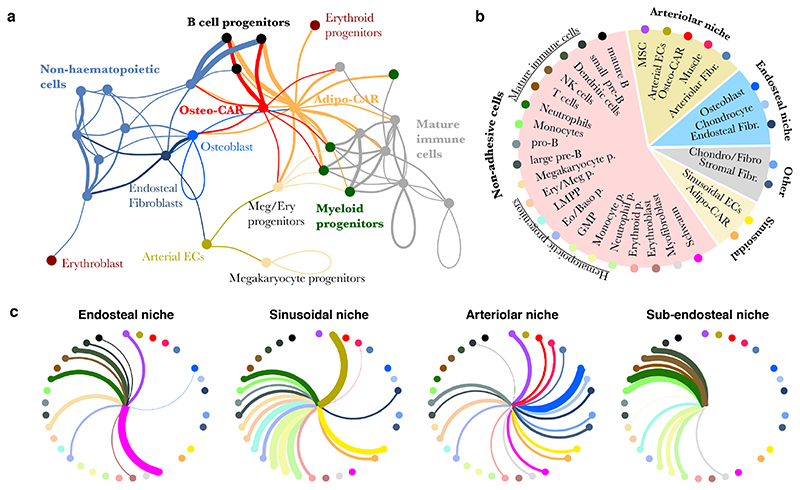
The bone marrow constitutes the primary site for life-long blood production and skeletal regeneration. However, its cellular and spatial organization remains controversial. Here, we combine single-cell and spatially resolved transcriptomics to systematically map the molecular, cellular and spatial composition of distinct bone marrow niches. This allowed us to transcriptionally profile all major bone-marrow-resident cell types, determine their localization and clarify sources of pro-haematopoietic factors. Our data demonstrate that Cxcl12-abundant-reticular (CAR) cell subsets (Adipo-CAR and Osteo-CAR) differentially localize to sinusoidal and arteriolar surfaces, act locally as 'professional cytokine-secreting cells' and thereby establish peri-vascular micro-niches. Importantly, the three-dimensional bone-marrow organization can be accurately inferred from single-cell transcriptome data using the RNA-Magnet algorithm described here. Together, our study reveals the cellular and spatial organization of bone marrow niches and offers a systematic approach to dissect the complex organization of whole organs.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Unraveling bone marrow architecture.Nat Cell Biol. 2020 Jan;22(1):5-6. doi: 10.1038/s41556-019-0447-6. Nat Cell Biol. 2020. PMID: 31871318 No abstract available.
References
-
- Ramasamy SK, et al. Regulation of Hematopoiesis and Osteogenesis by Blood Vessel-Derived Signals. Annu Rev Cell Dev Biol. 2016;32:649–675. - PubMed
-
- Sugiyama T, Kohara H, Noda M, Nagasawa T. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity. 2006;25:977–88. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
