

Sideroflexin 4 affects Fe-S cluster biogenesis, iron metabolism, mitochondrial respiration and heme biosynthetic enzymes
- PMID: 31873120
- PMCID: PMC6928202
- DOI: 10.1038/s41598-019-55907-z
Sideroflexin 4 affects Fe-S cluster biogenesis, iron metabolism, mitochondrial respiration and heme biosynthetic enzymes
Abstract
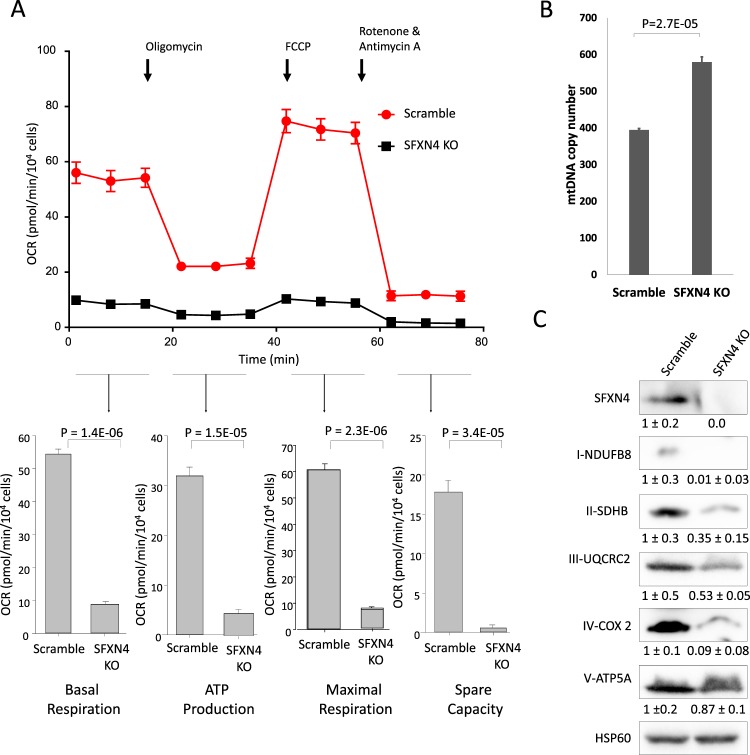
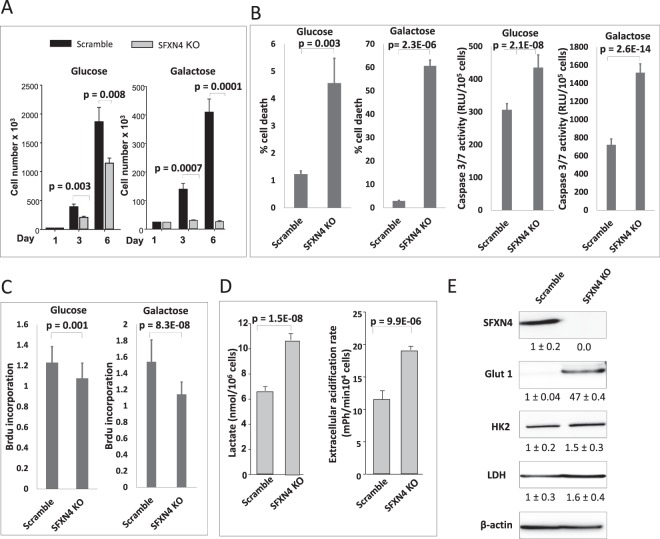
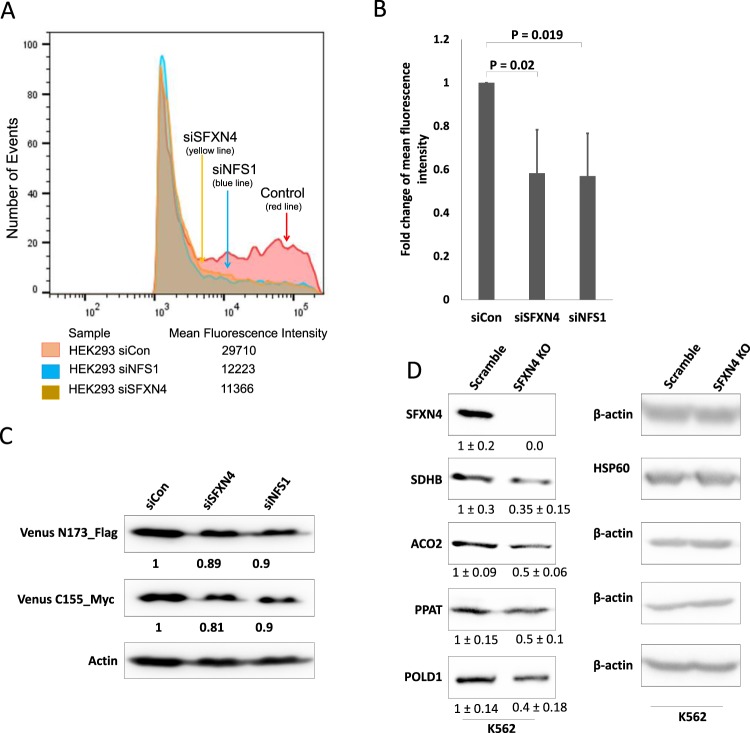
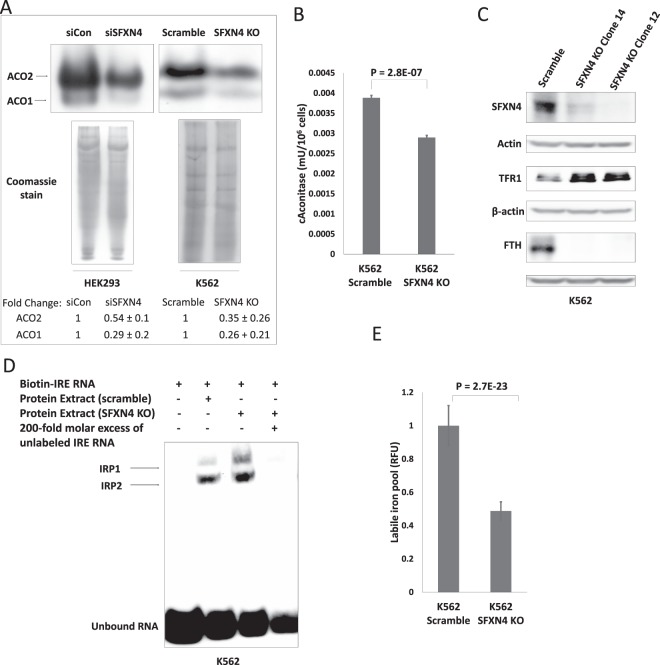
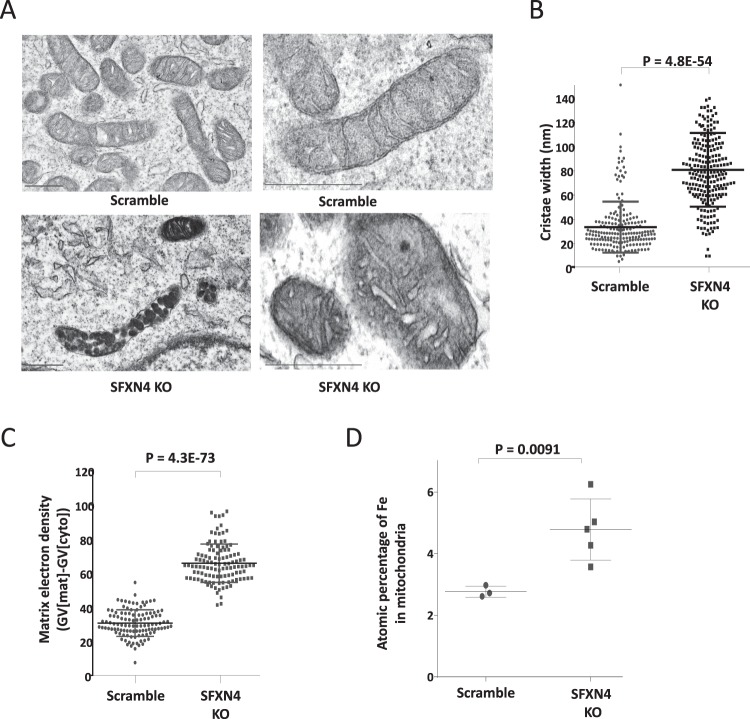
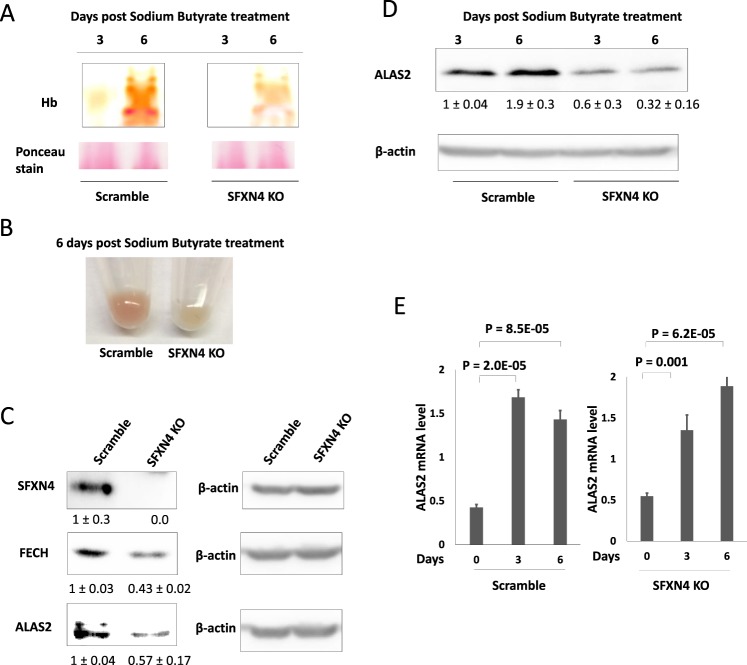
Sideroflexin4 (SFXN4) is a member of a family of nuclear-encoded mitochondrial proteins. Rare germline mutations in SFXN4 lead to phenotypic characteristics of mitochondrial disease including impaired mitochondrial respiration and hematopoetic abnormalities. We sought to explore the function of this protein. We show that knockout of SFXN4 has profound effects on Fe-S cluster formation. This in turn diminishes mitochondrial respiratory chain complexes and mitochondrial respiration and causes a shift to glycolytic metabolism. SFXN4 knockdown reduces the stability and activity of cellular Fe-S proteins, affects iron metabolism by influencing the cytosolic aconitase-IRP1 switch, redistributes iron from the cytosol to mitochondria, and impacts heme synthesis by reducing levels of ferrochelatase and inhibiting translation of ALAS2. We conclude that SFXN4 is essential for normal functioning of mitochondria, is necessary for Fe-S cluster biogenesis and iron homeostasis, and plays a critical role in mitochondrial respiration and synthesis of heme.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
