

Autism risk in offspring can be assessed through quantification of male sperm mosaicism
- PMID: 31873310
- PMCID: PMC7032648
- DOI: 10.1038/s41591-019-0711-0
Autism risk in offspring can be assessed through quantification of male sperm mosaicism
Abstract
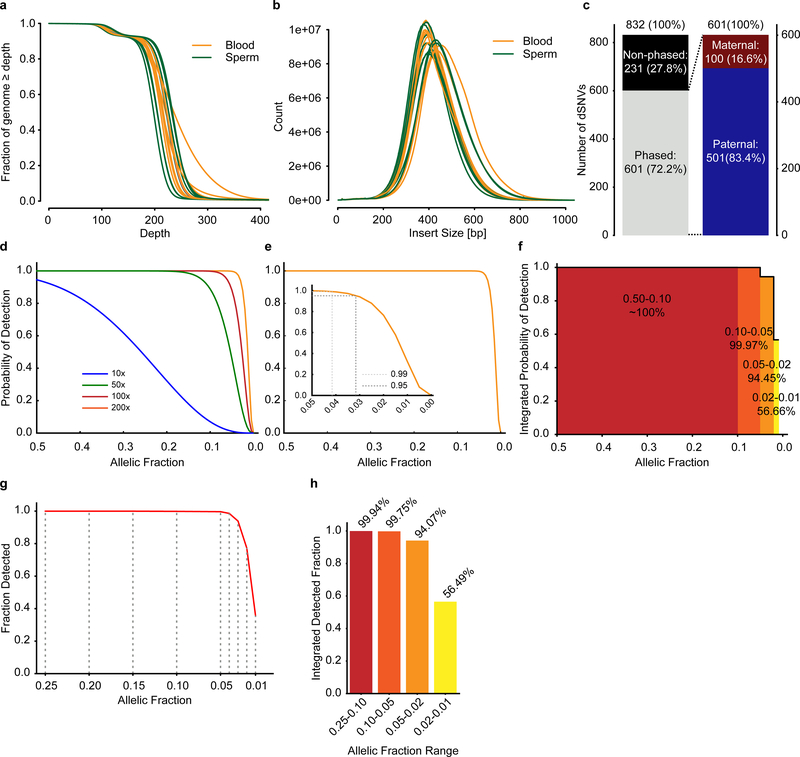
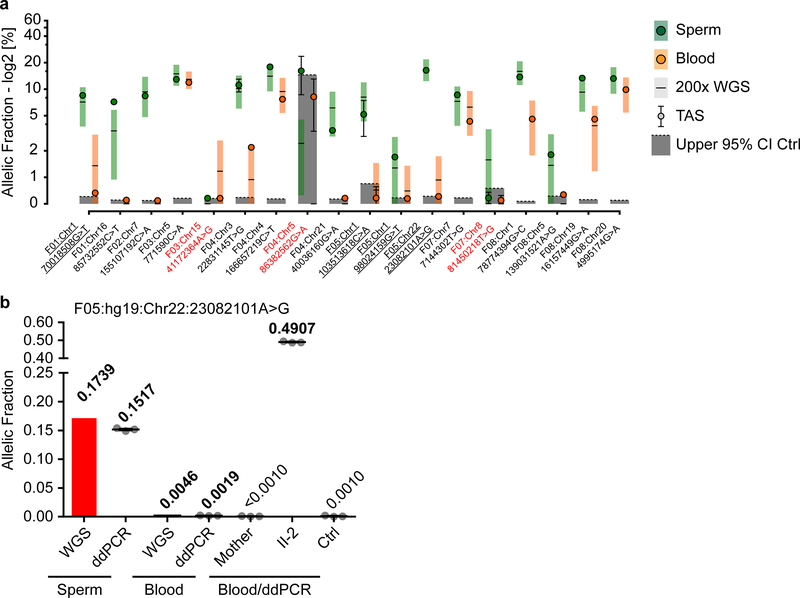
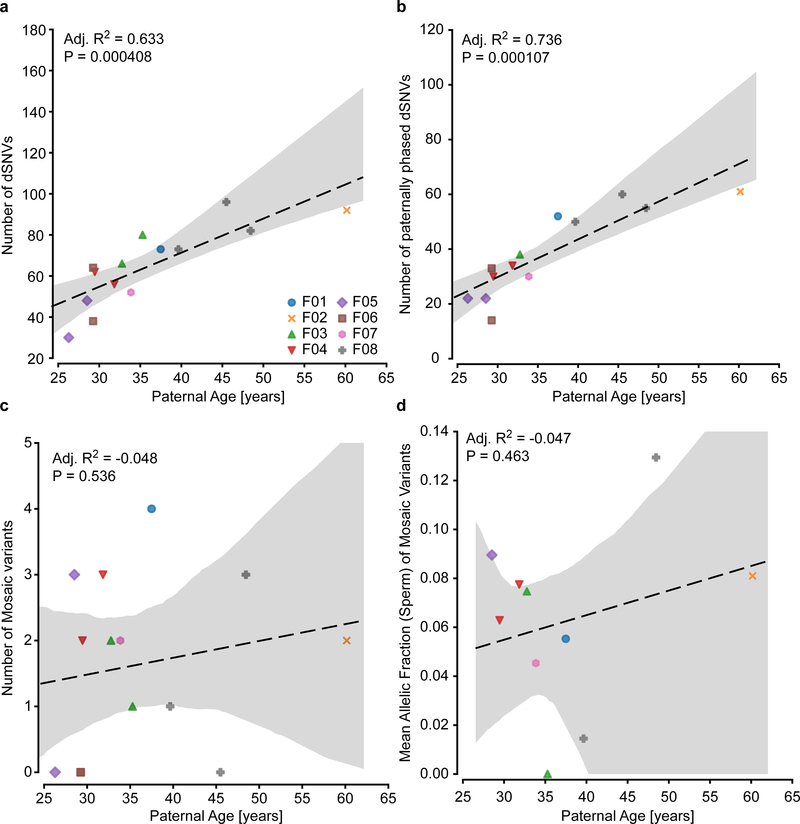
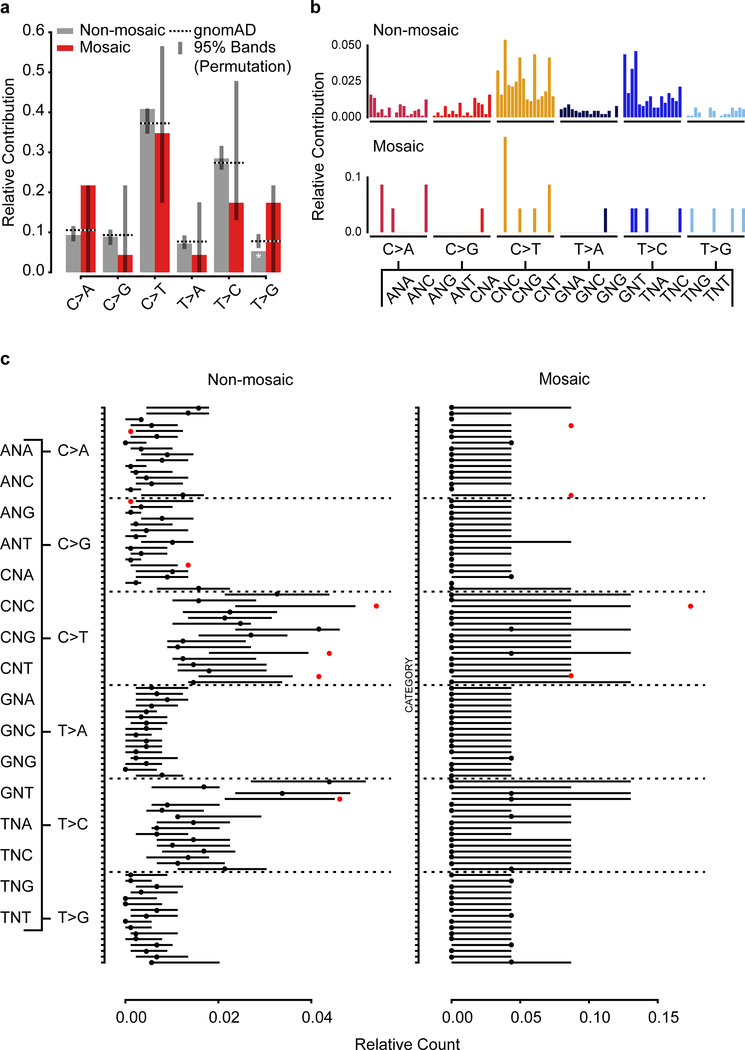
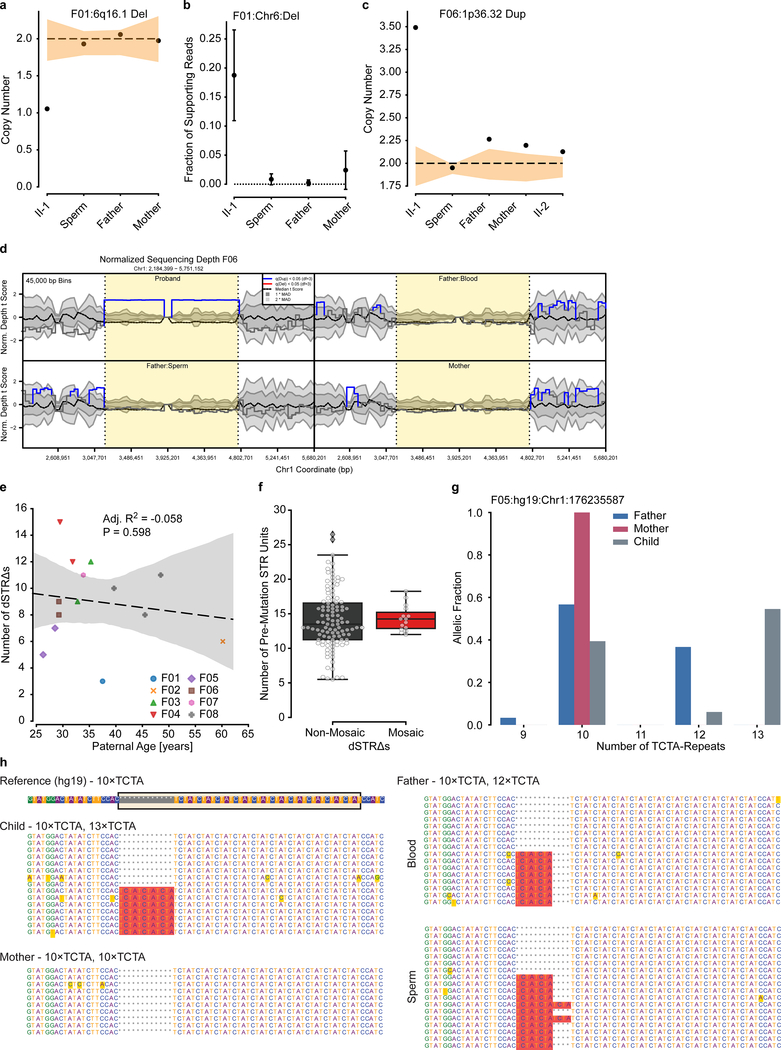
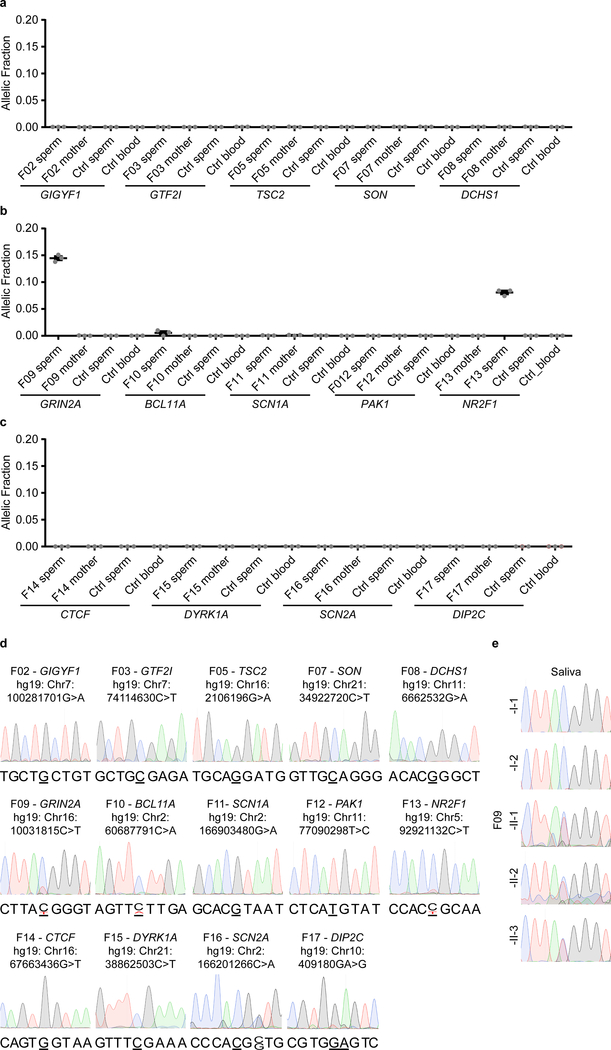
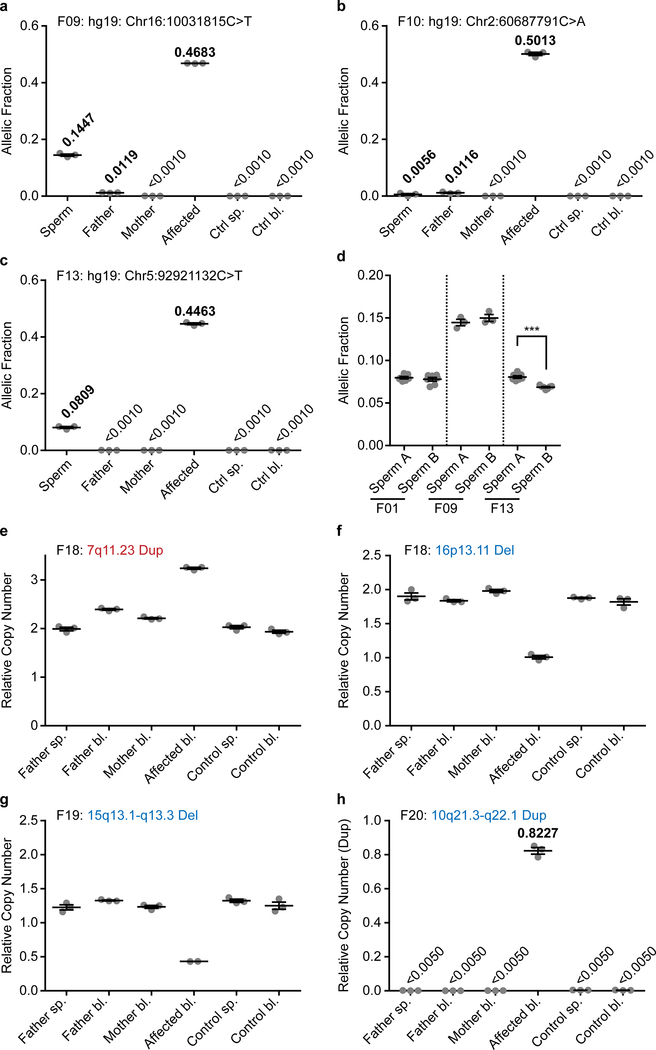
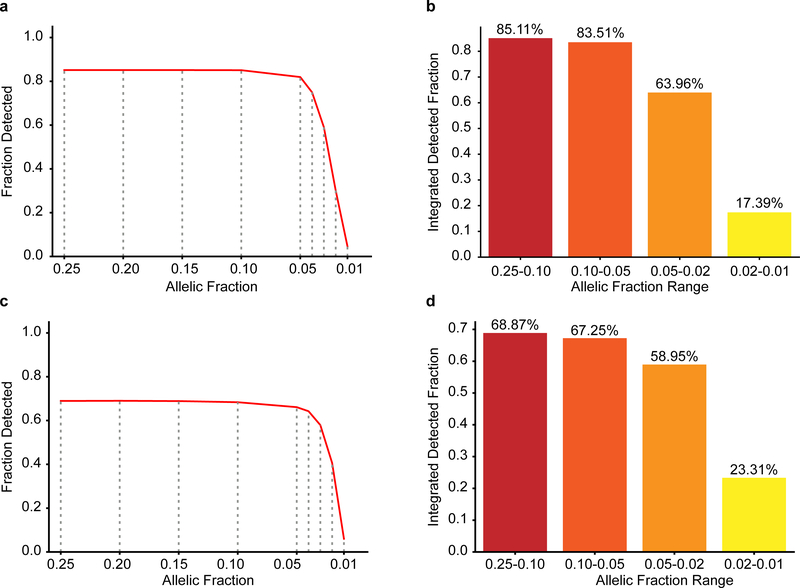
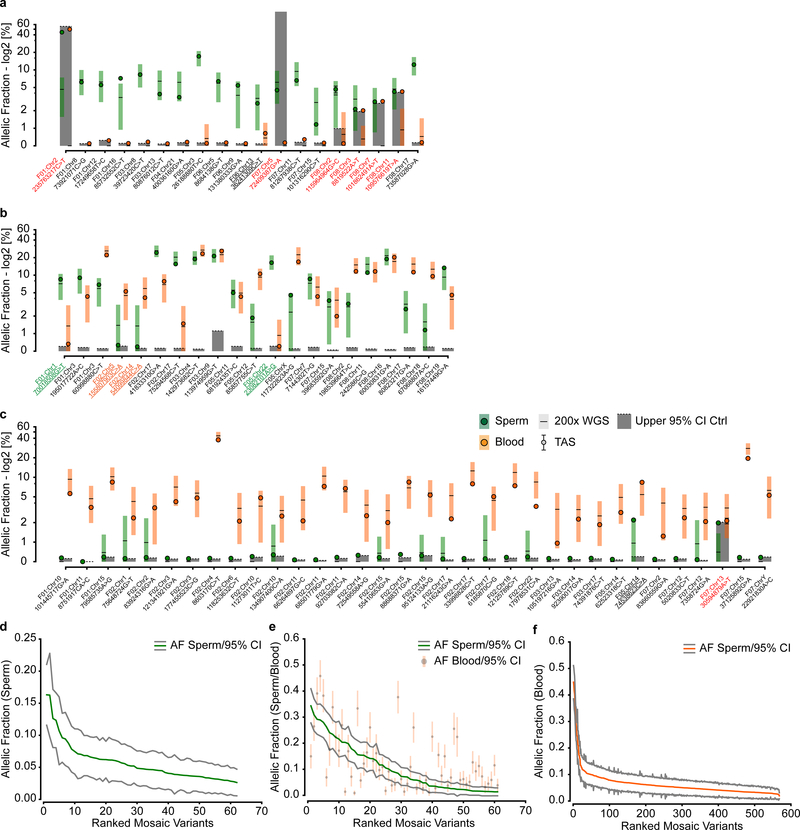
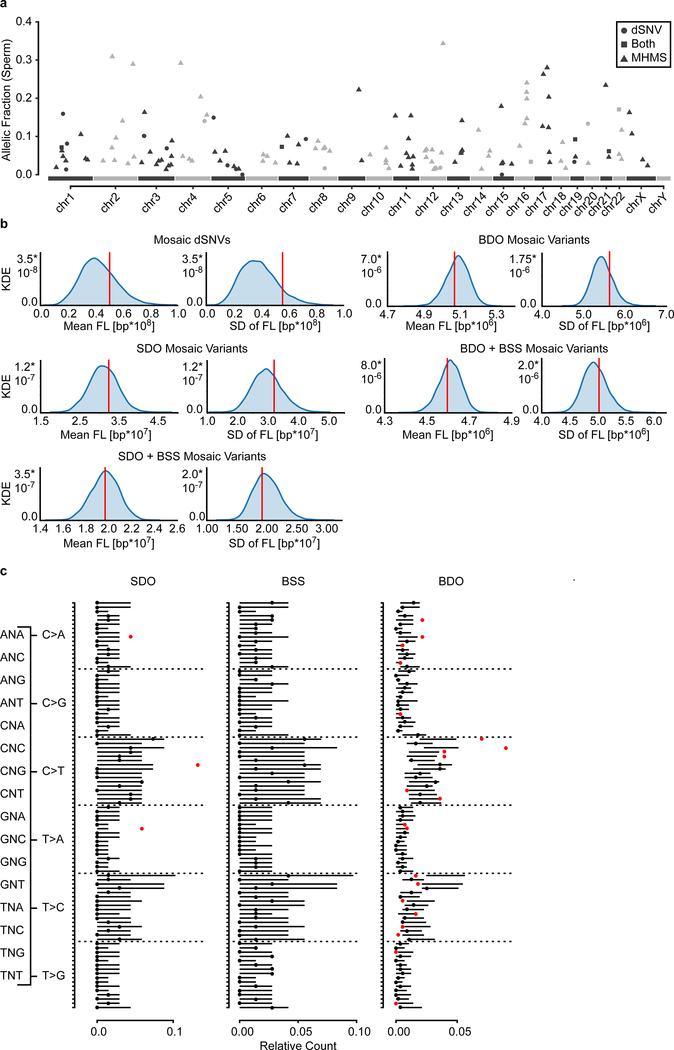
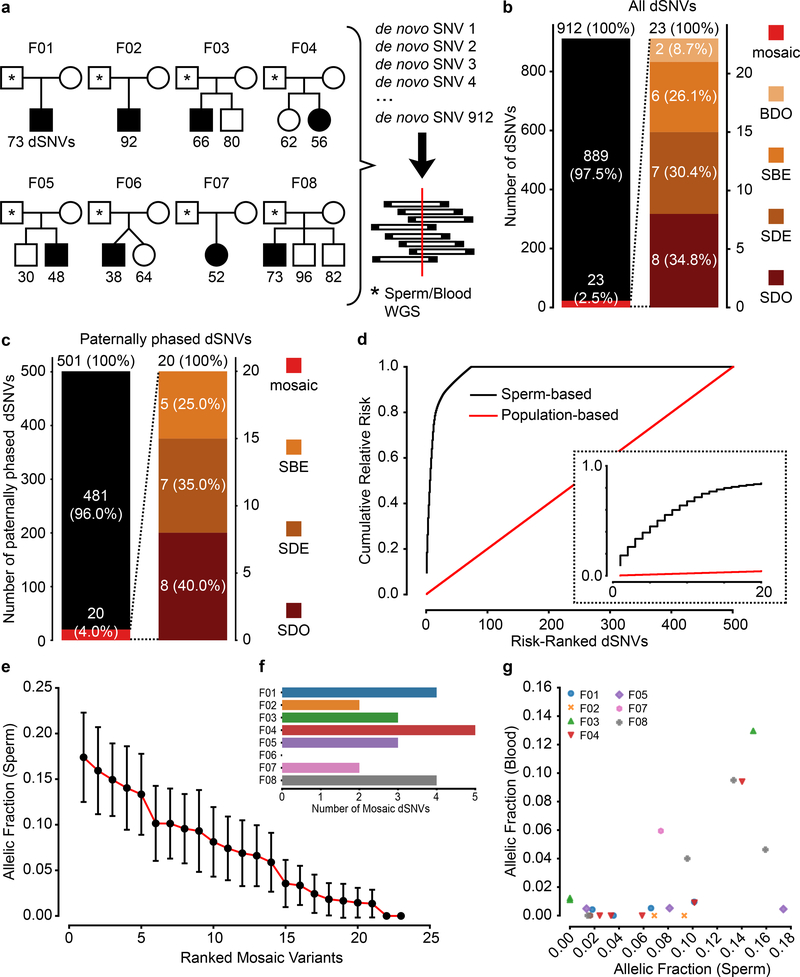
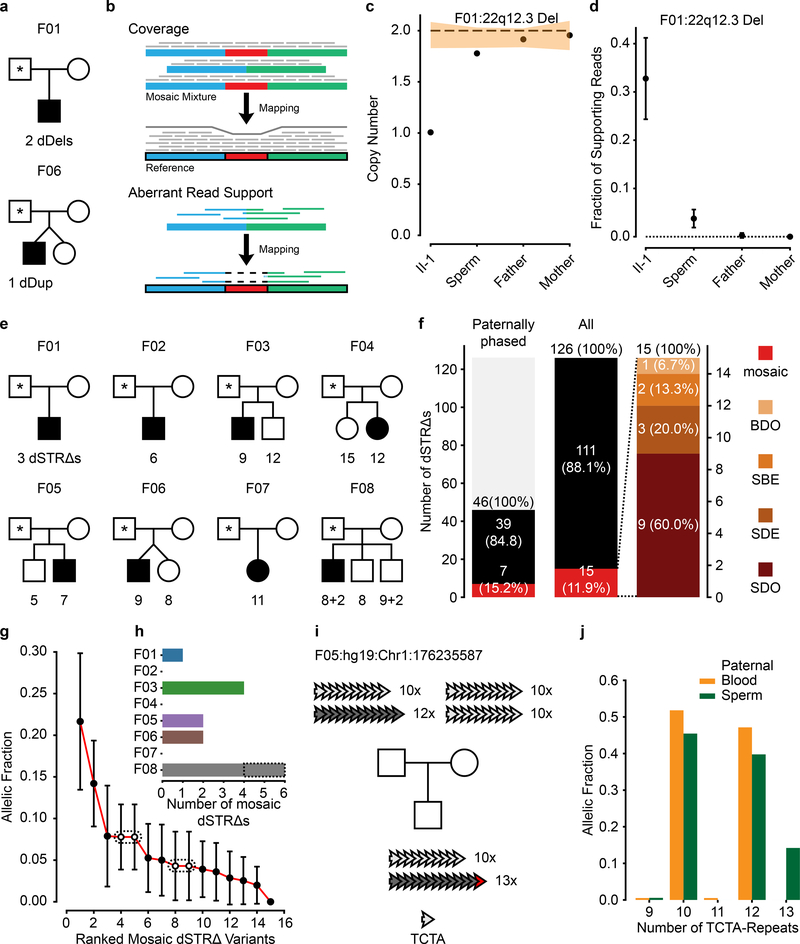
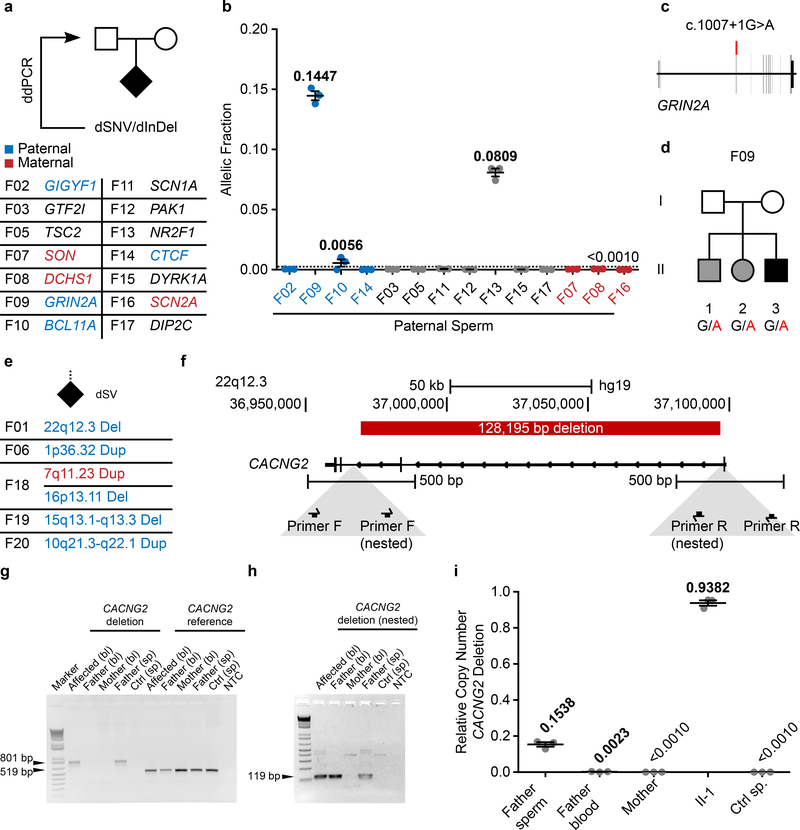
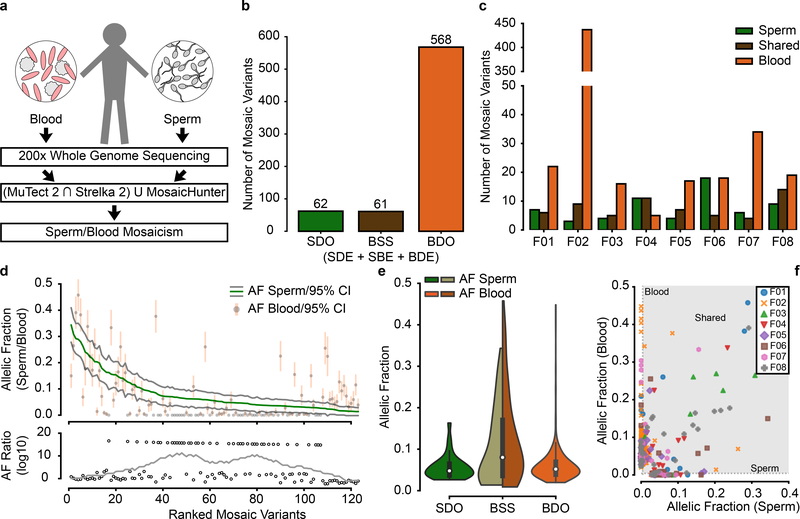
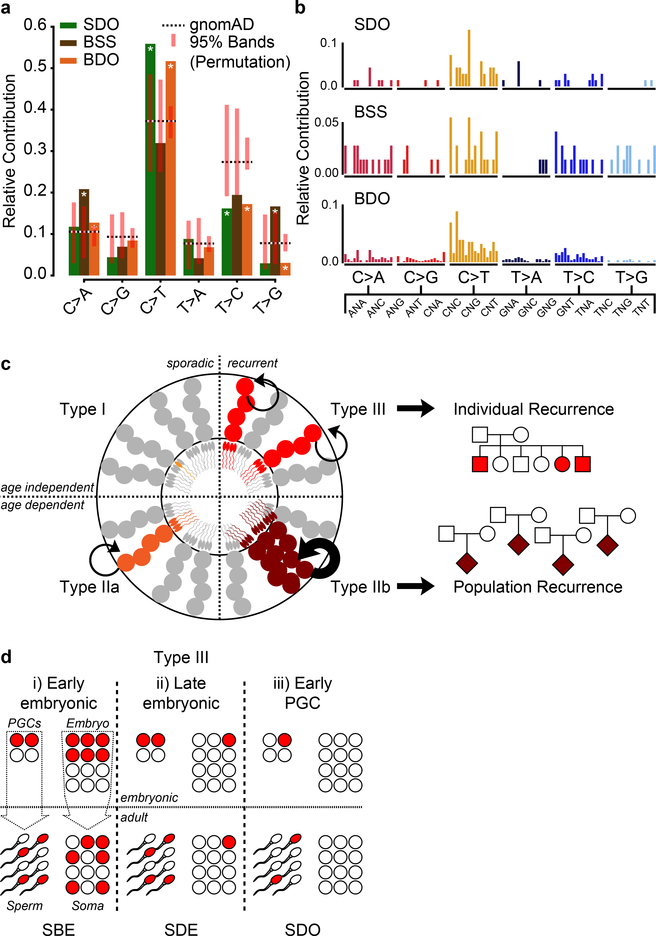
De novo mutations arising on the paternal chromosome make the largest known contribution to autism risk, and correlate with paternal age at the time of conception. The recurrence risk for autism spectrum disorders is substantial, leading many families to decline future pregnancies, but the potential impact of assessing parental gonadal mosaicism has not been considered. We measured sperm mosaicism using deep-whole-genome sequencing, for variants both present in an offspring and evident only in father's sperm, and identified single-nucleotide, structural and short tandem-repeat variants. We found that mosaicism quantification can stratify autism spectrum disorders recurrence risk due to de novo mutations into a vast majority with near 0% recurrence and a small fraction with a substantially higher and quantifiable risk, and we identify novel mosaic variants at risk for transmission to a future offspring. This suggests, therefore, that genetic counseling would benefit from the addition of sperm mosaicism assessment.
Figures
Comment in
-
Paternal sperm DNA mosaicism and recurrence risk of autism in families.Nat Med. 2020 Jan;26(1):26-28. doi: 10.1038/s41591-019-0732-8. Nat Med. 2020. PMID: 31873313 No abstract available.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
