

Microbial bile acid metabolites modulate gut RORγ+ regulatory T cell homeostasis
- PMID: 31875848
- PMCID: PMC7274525
- DOI: 10.1038/s41586-019-1865-0
Microbial bile acid metabolites modulate gut RORγ+ regulatory T cell homeostasis
Abstract
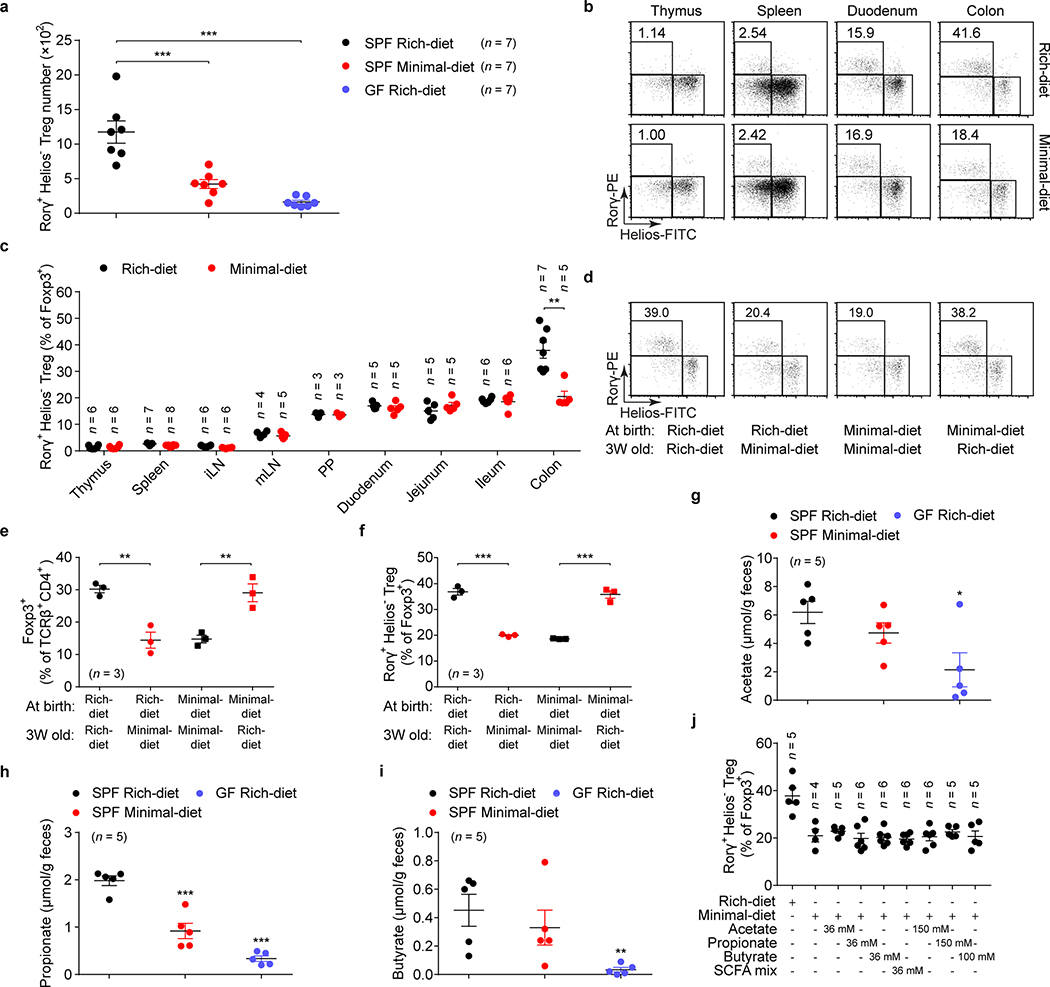
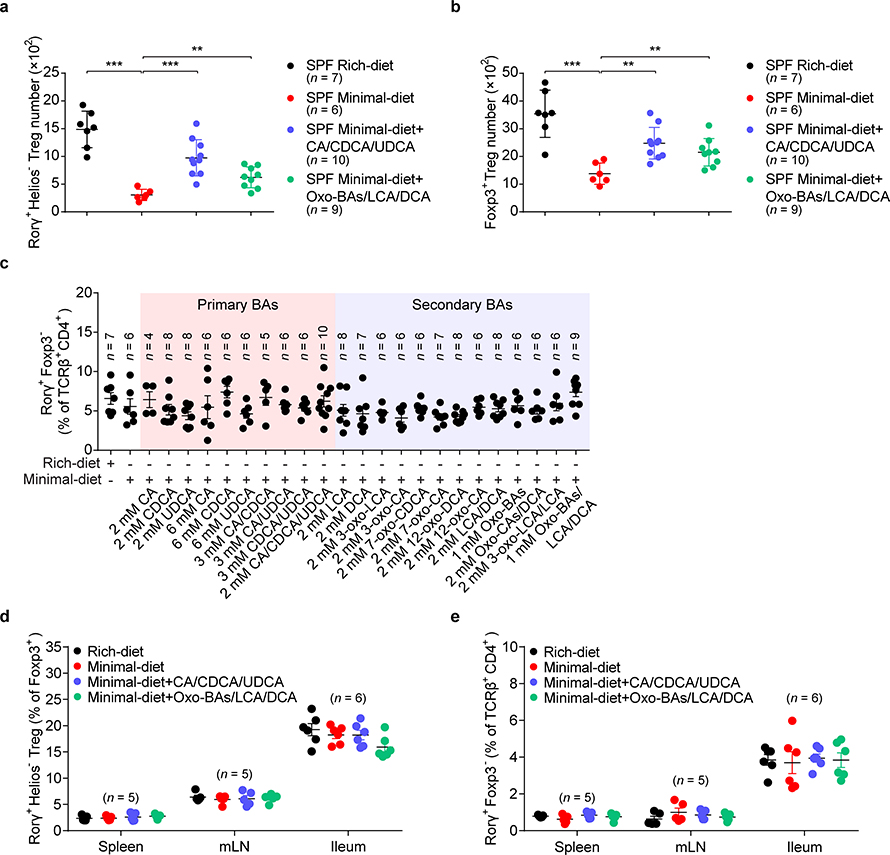
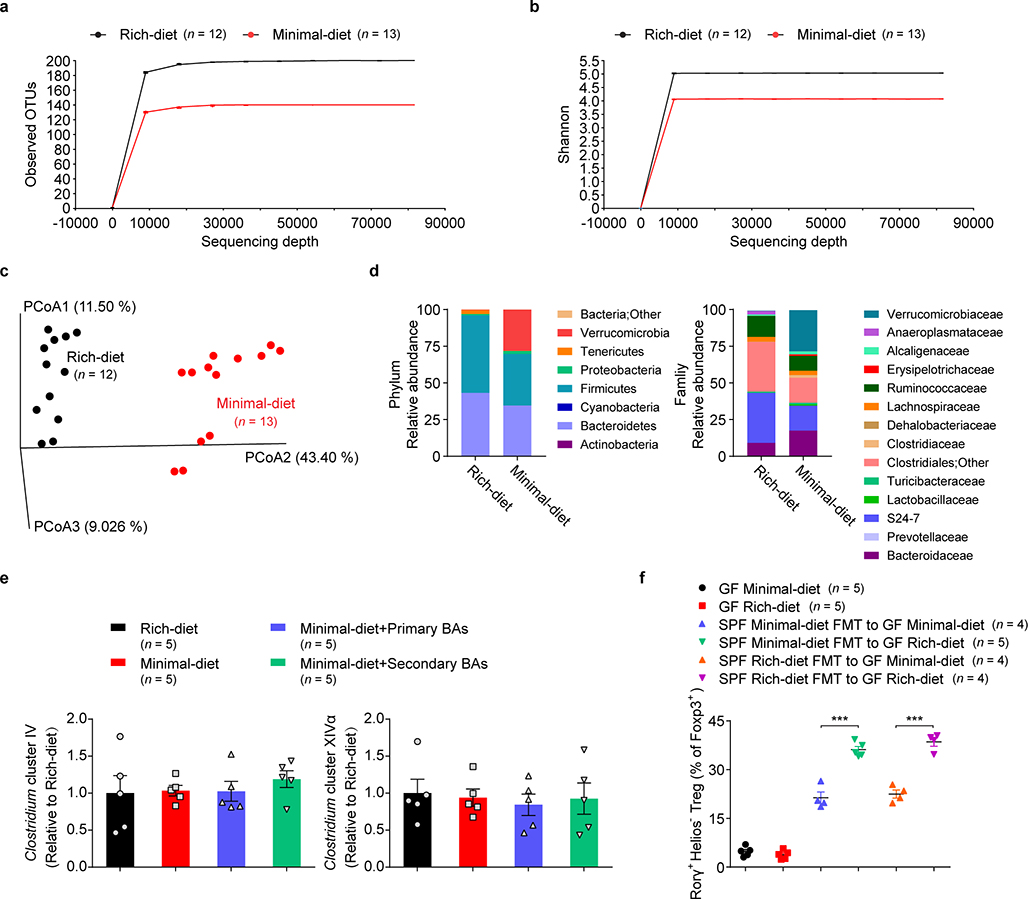
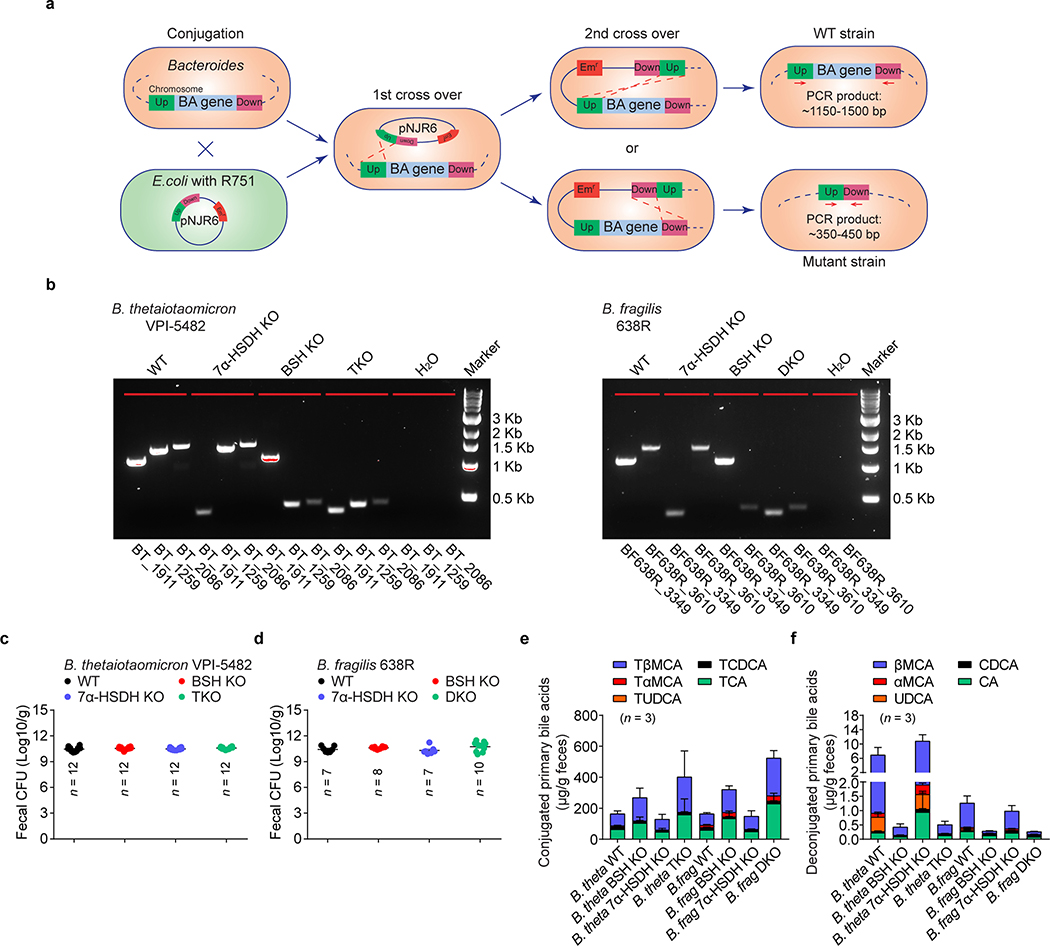
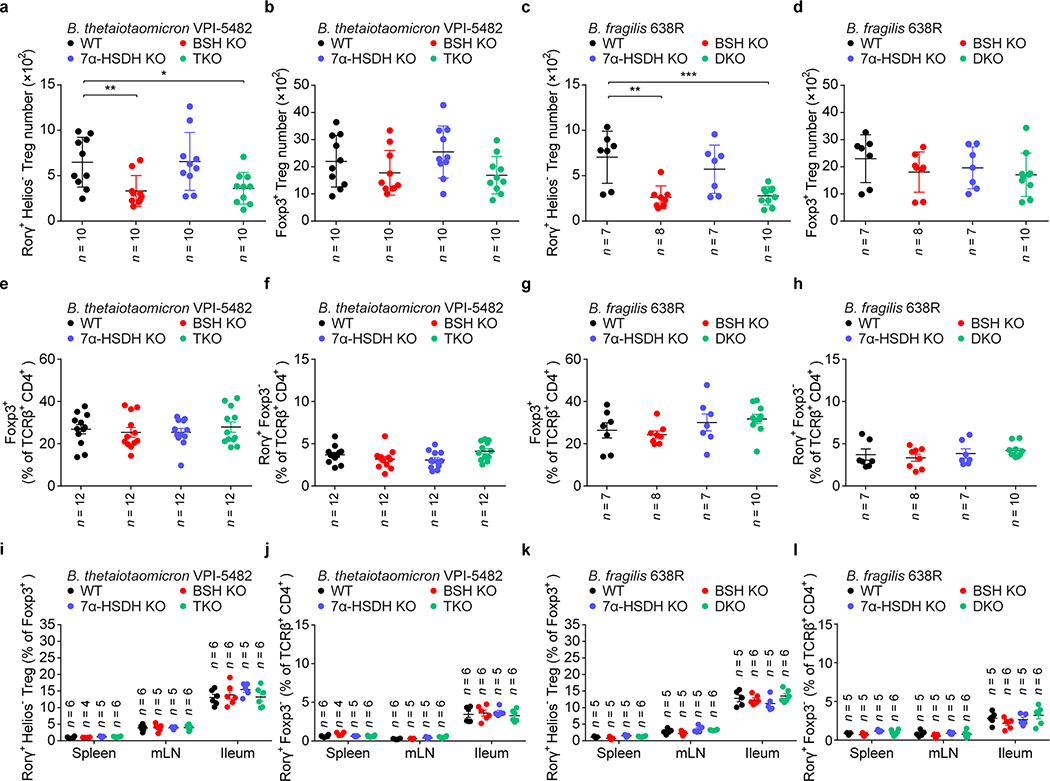
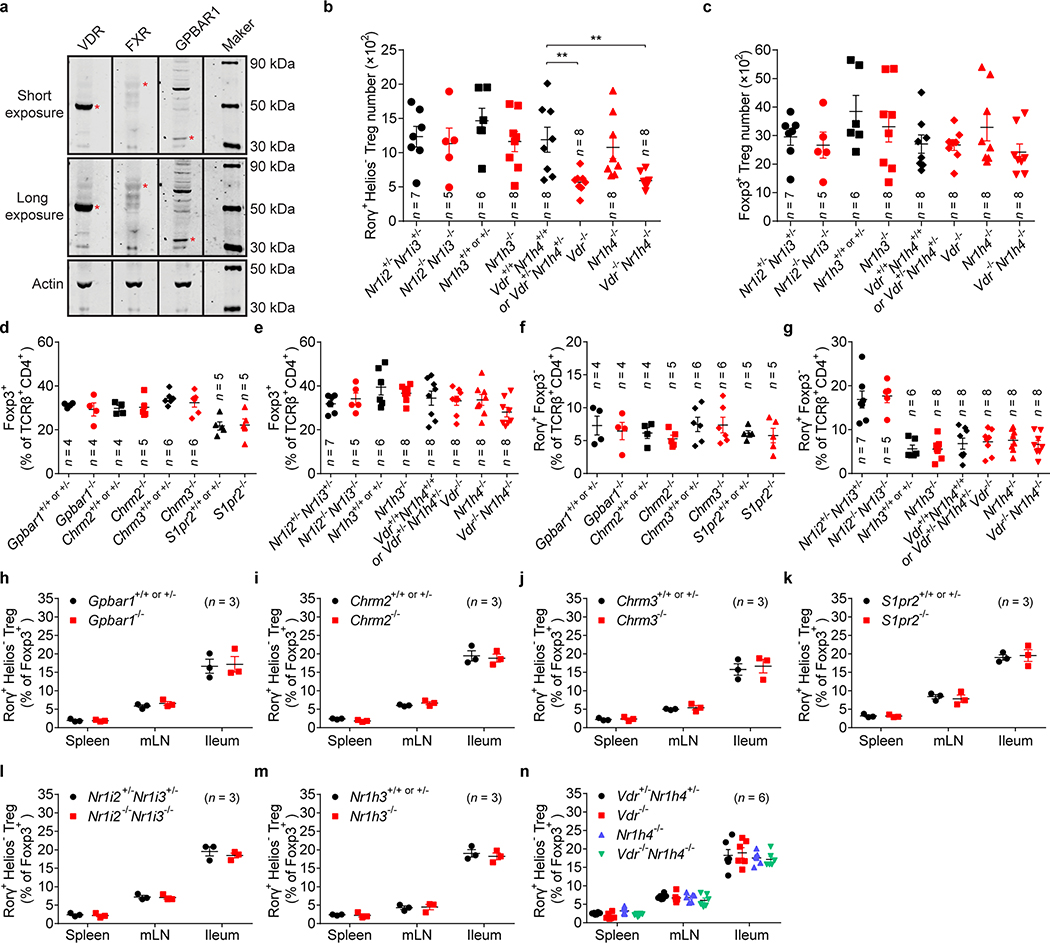
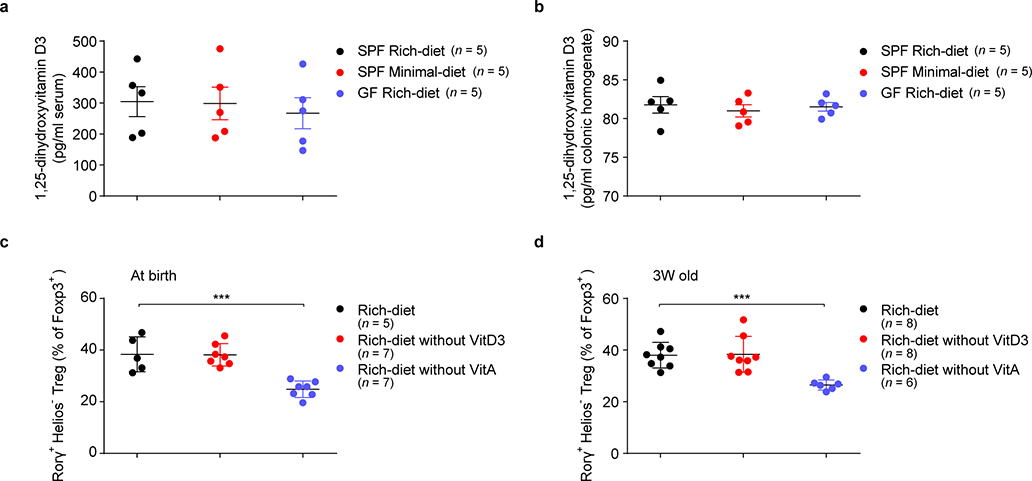
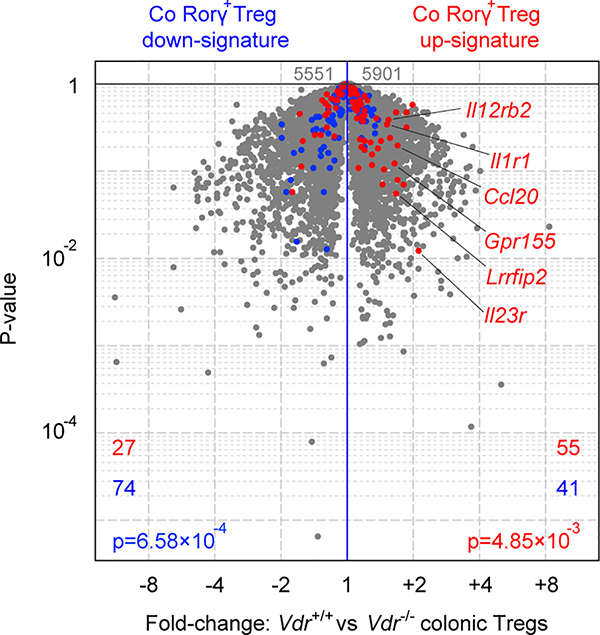
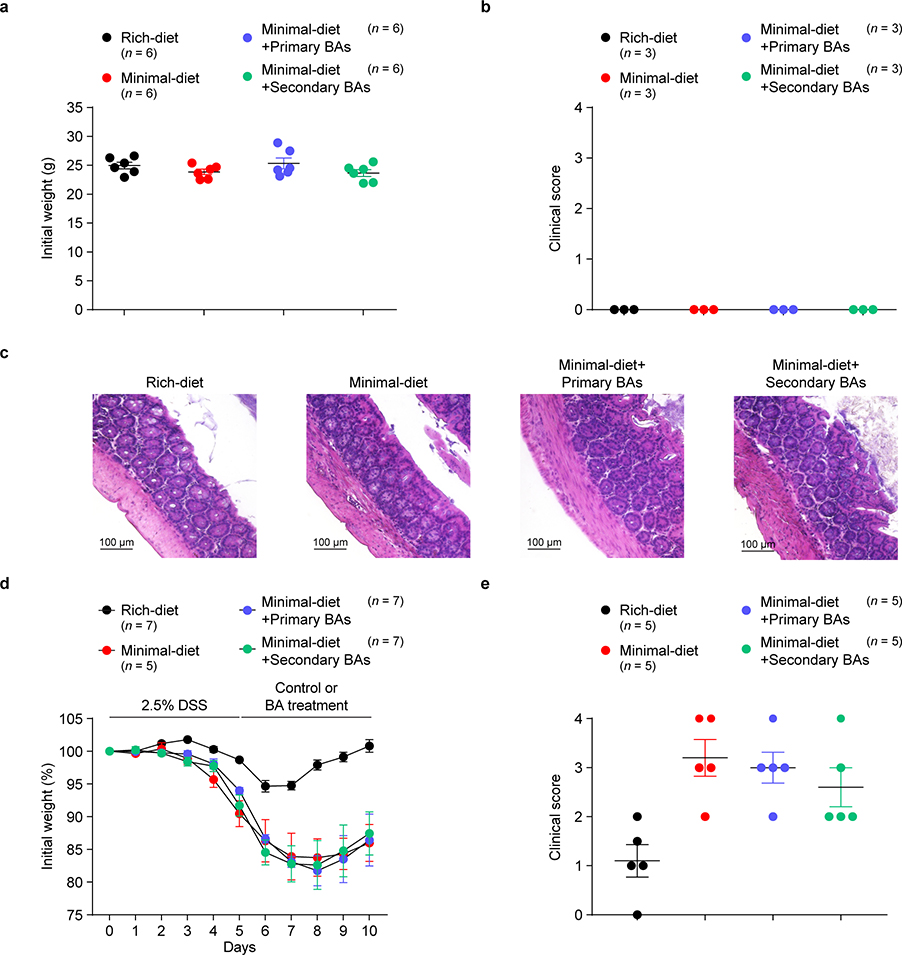
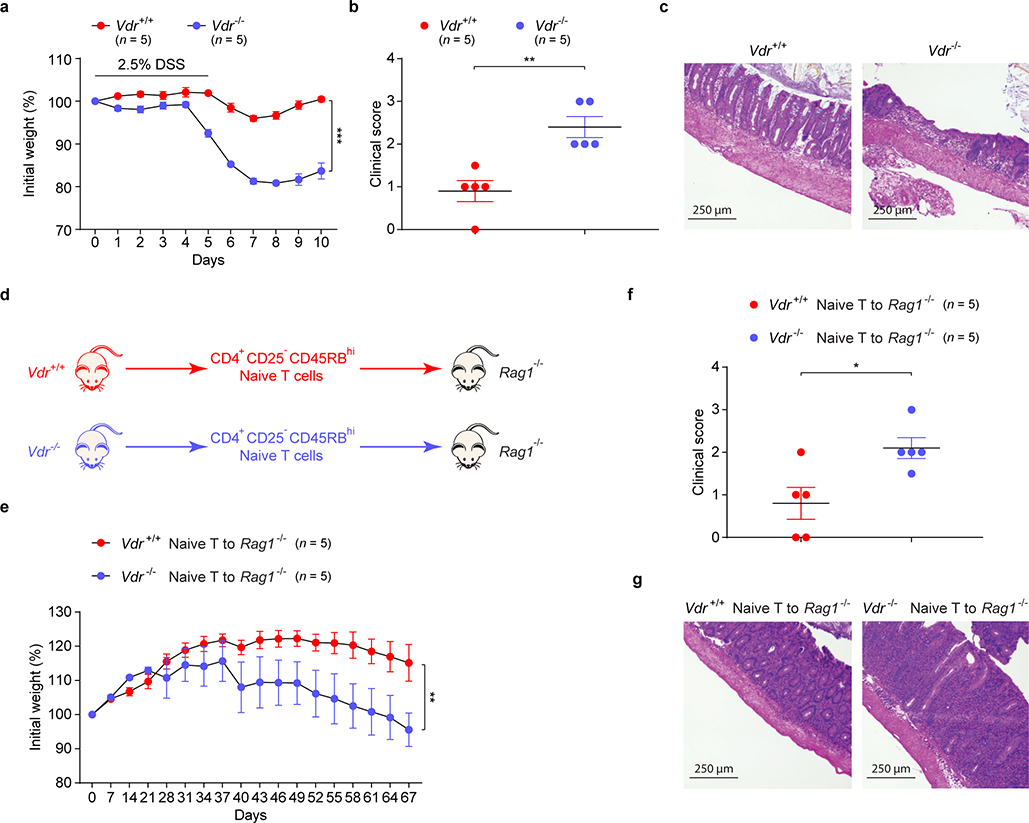
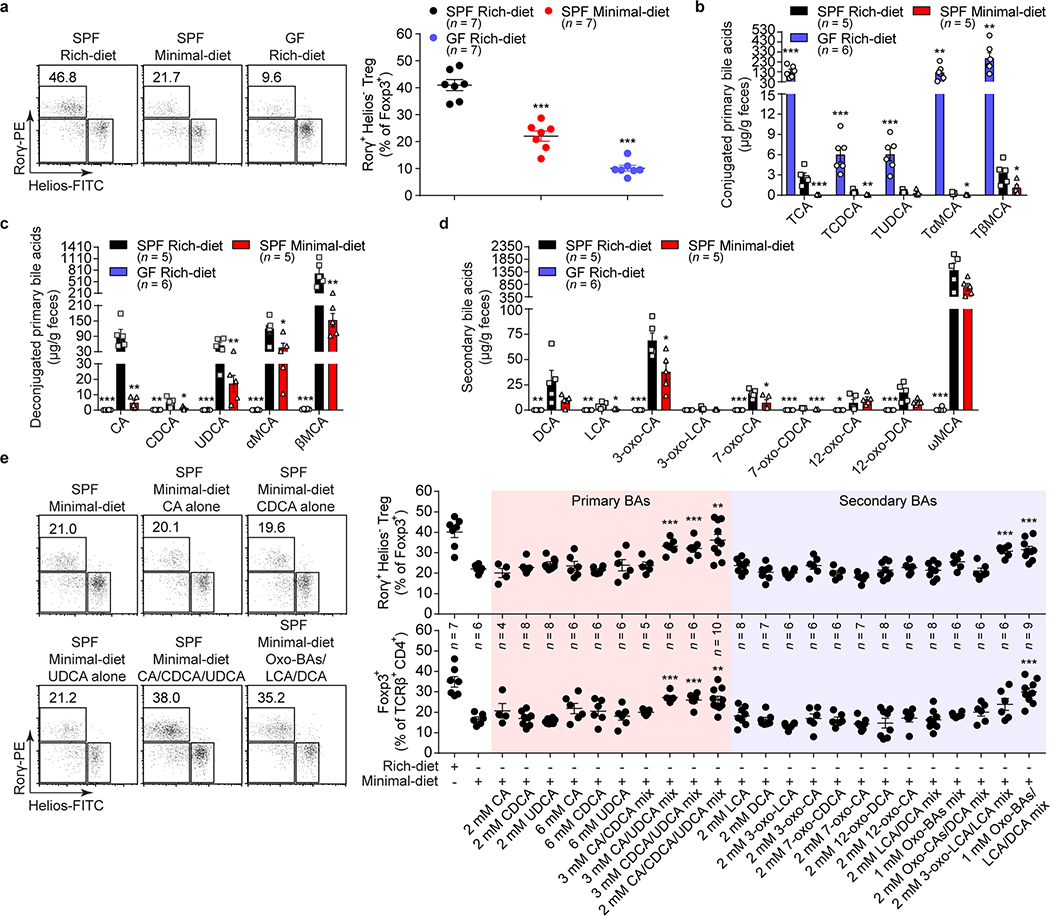
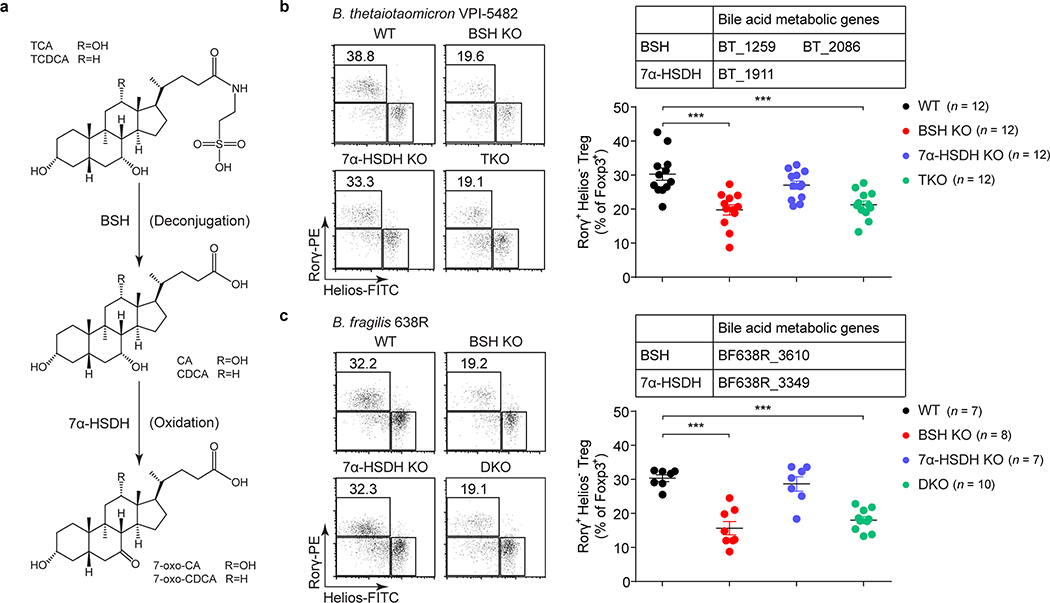
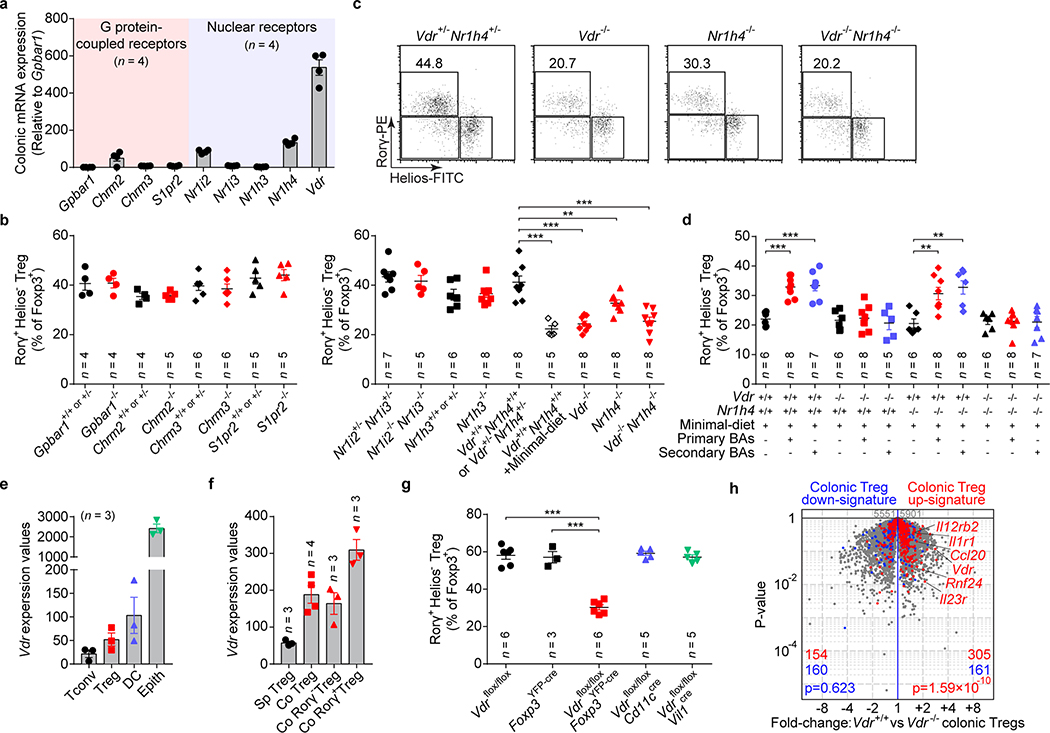
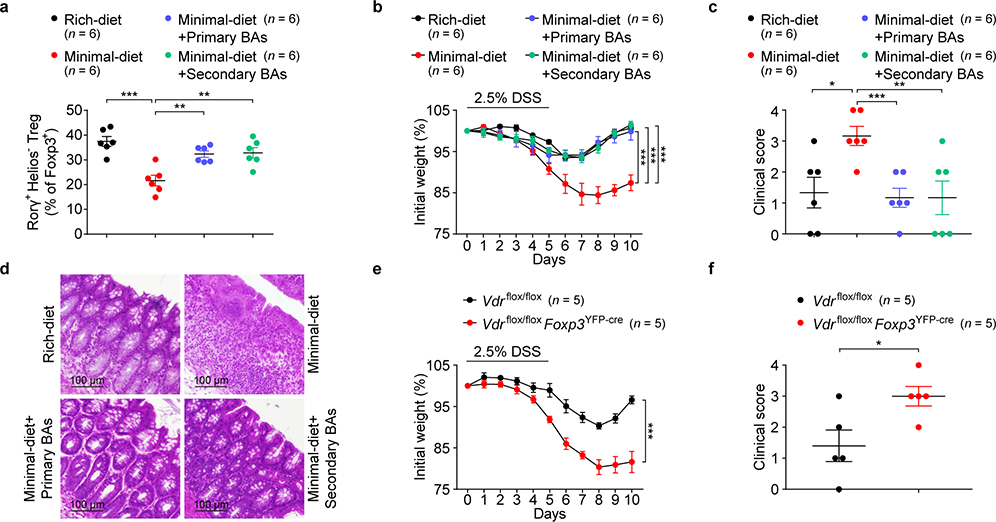
The metabolic pathways encoded by the human gut microbiome constantly interact with host gene products through numerous bioactive molecules1. Primary bile acids (BAs) are synthesized within hepatocytes and released into the duodenum to facilitate absorption of lipids or fat-soluble vitamins2. Some BAs (approximately 5%) escape into the colon, where gut commensal bacteria convert them into various intestinal BAs2 that are important hormones that regulate host cholesterol metabolism and energy balance via several nuclear receptors and/or G-protein-coupled receptors3,4. These receptors have pivotal roles in shaping host innate immune responses1,5. However, the effect of this host-microorganism biliary network on the adaptive immune system remains poorly characterized. Here we report that both dietary and microbial factors influence the composition of the gut BA pool and modulate an important population of colonic FOXP3+ regulatory T (Treg) cells expressing the transcription factor RORγ. Genetic abolition of BA metabolic pathways in individual gut symbionts significantly decreases this Treg cell population. Restoration of the intestinal BA pool increases colonic RORγ+ Treg cell counts and ameliorates host susceptibility to inflammatory colitis via BA nuclear receptors. Thus, a pan-genomic biliary network interaction between hosts and their bacterial symbionts can control host immunological homeostasis via the resulting metabolites.
Conflict of interest statement
Competing Interests
The authors declare no competing interests.
Figures
Comment in
-
Microbiome Modulation of the Host Adaptive Immunity through Bile Acid Modification.Cell Metab. 2020 Mar 3;31(3):445-447. doi: 10.1016/j.cmet.2020.02.006. Cell Metab. 2020. PMID: 32130878
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
