

Type IV CRISPR-Cas systems are highly diverse and involved in competition between plasmids
- PMID: 31879772
- PMCID: PMC7038947
- DOI: 10.1093/nar/gkz1197
Type IV CRISPR-Cas systems are highly diverse and involved in competition between plasmids
Abstract
CRISPR-Cas systems provide prokaryotes with adaptive immune functions against viruses and other genetic parasites. In contrast to all other types of CRISPR-Cas systems, type IV has remained largely overlooked. Here, we describe a previously uncharted diversity of type IV gene cassettes, primarily encoded by plasmid-like elements from diverse prokaryotic taxa. Remarkably, via a comprehensive analysis of their CRISPR spacer content, these systems were found to exhibit a strong bias towards the targeting of other plasmids. Our data indicate that the functions of type IV systems have diverged from those of other host-related CRISPR-Cas immune systems to adopt a role in mediating conflicts between plasmids. Furthermore, we find evidence for cross-talk between certain type IV and type I CRISPR-Cas systems that co-exist intracellularly, thus providing a simple answer to the enigmatic absence of type IV adaptation modules. Collectively, our results lead to the expansion and reclassification of type IV systems and provide novel insights into the biological function and evolution of these elusive systems.
© The Author(s) 2019. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
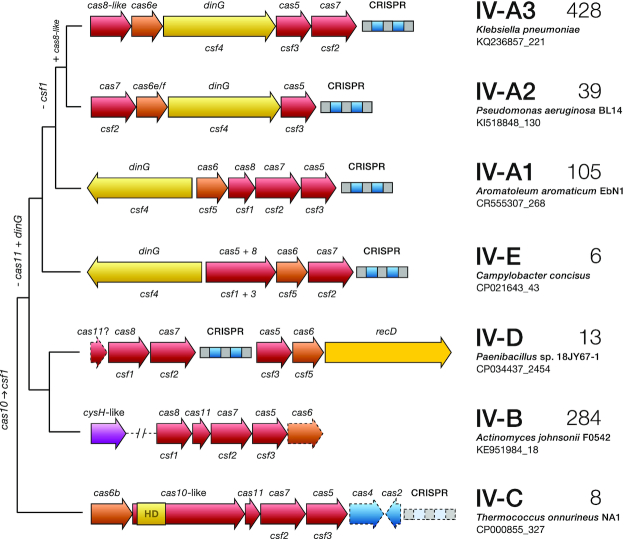
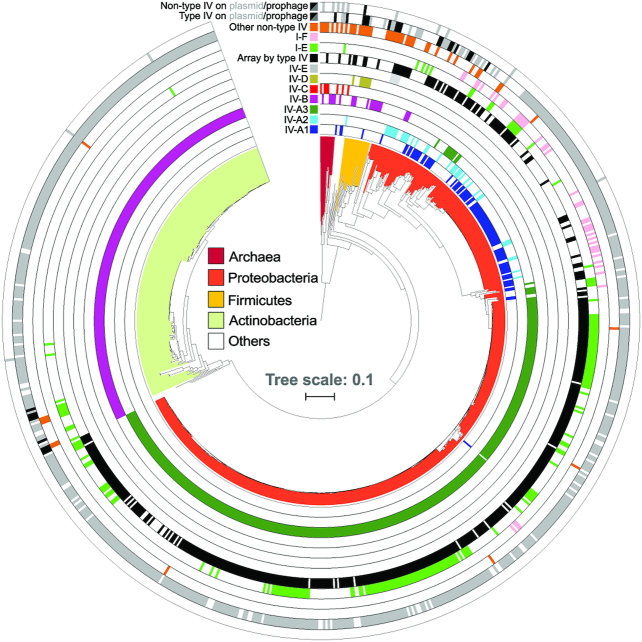
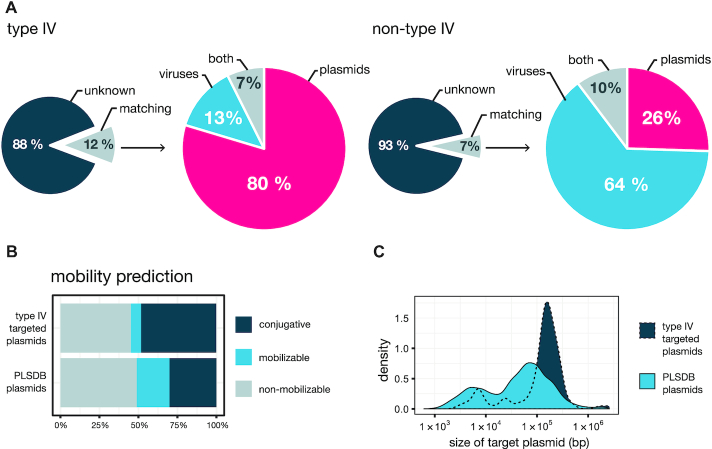
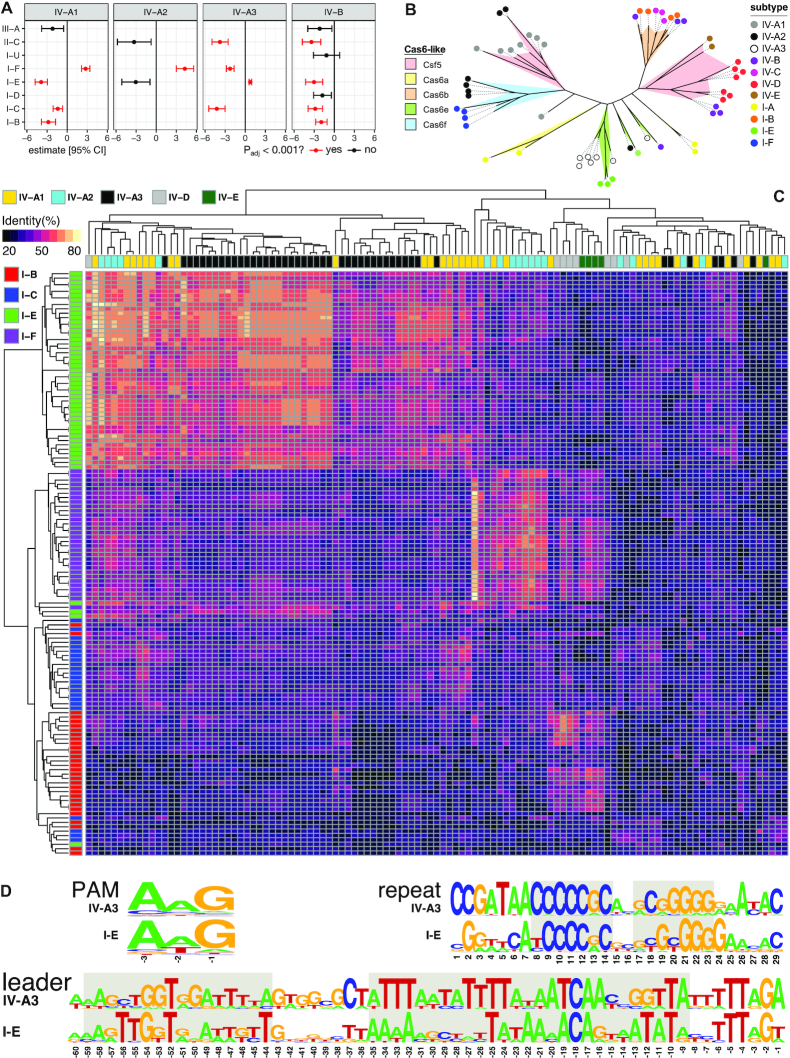
References
-
- Barrangou R., Doudna J.A.. Applications of CRISPR technologies in research and beyond. Nat. Biotechnol. 2016; 34:933–941. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
