

The structural features that distinguish PD-L2 from PD-L1 emerged in placental mammals
- PMID: 31882544
- PMCID: PMC7135984
- DOI: 10.1074/jbc.AC119.011747
The structural features that distinguish PD-L2 from PD-L1 emerged in placental mammals
Abstract
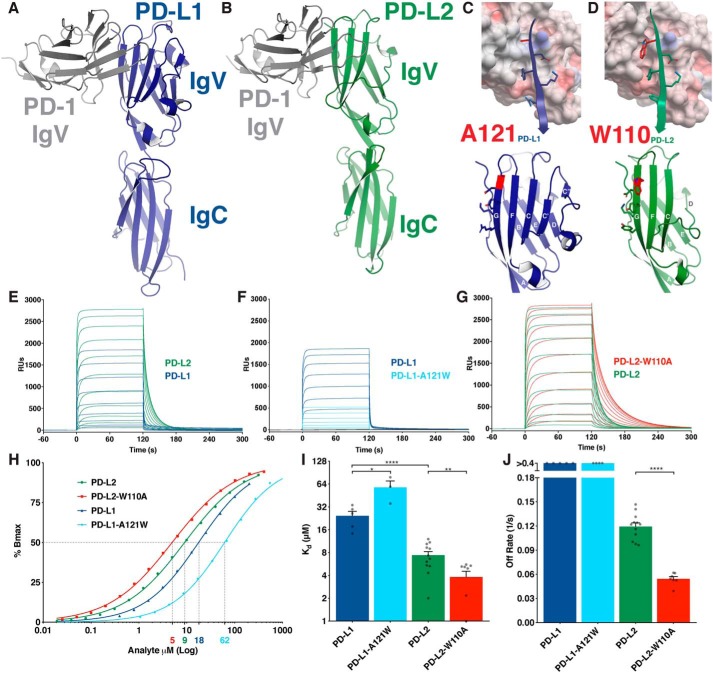
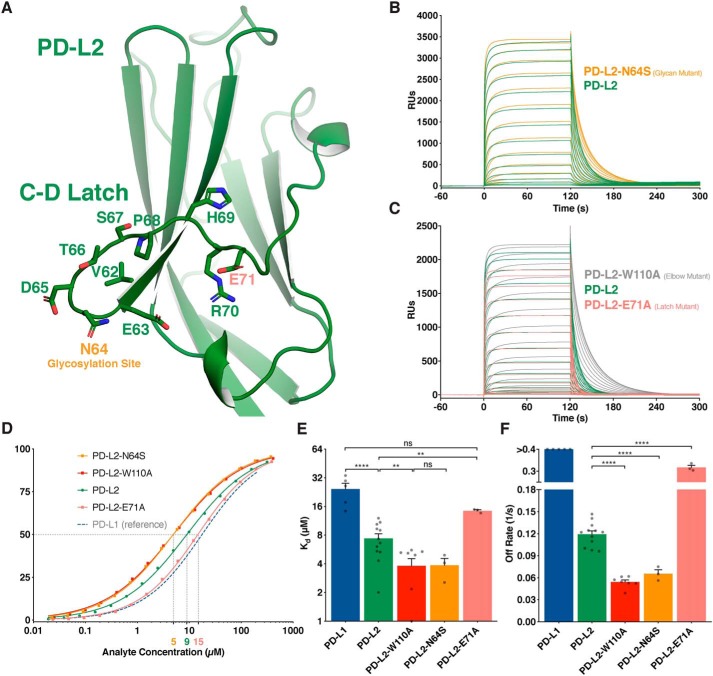
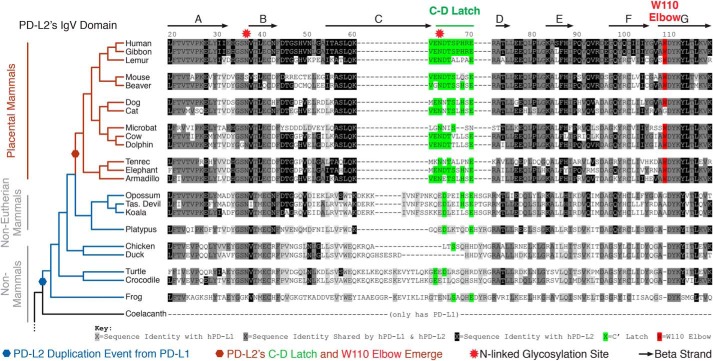
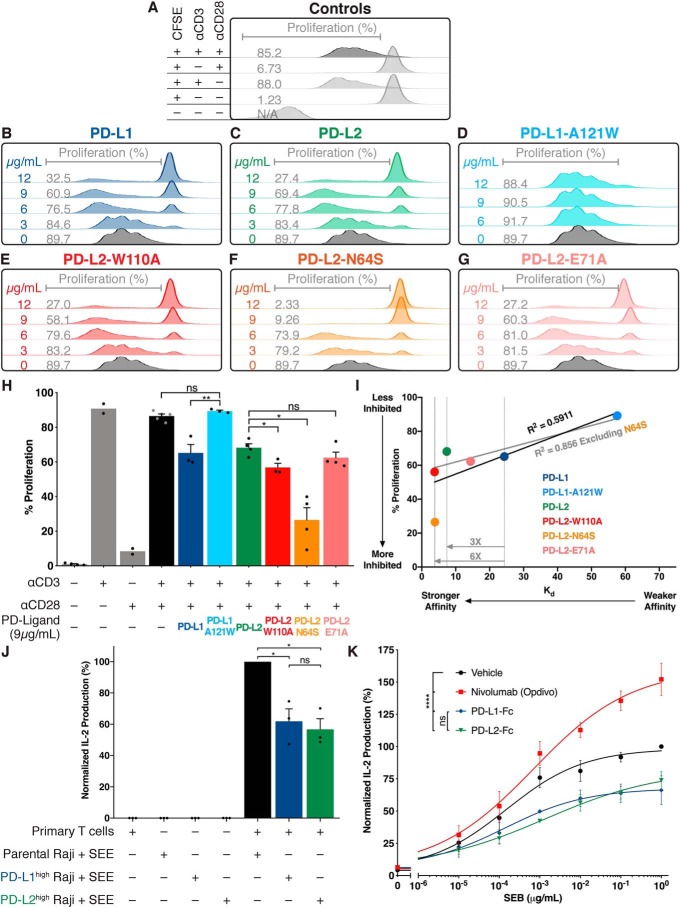
Programmed cell death protein 1 (PD-1) is an inhibitory receptor on T lymphocytes that is critical for modulating adaptive immunity. As such, it has been successfully exploited for cancer immunotherapy. Programmed death ligand 1 (PD-L1) and PD-L2 are ligands for PD-1; the former is ubiquitously expressed in inflamed tissues, whereas the latter is restricted to antigen-presenting cells. PD-L2 binds to PD-1 with 3-fold stronger affinity compared with PD-L1. To date, this affinity discrepancy has been attributed to a tryptophan (W110PD-L2) that is unique to PD-L2 and has been assumed to fit snuggly into a pocket on the PD-1 surface. Contrary to this model, using surface plasmon resonance to monitor real-time binding of recombinantly-expressed and -purified proteins, we found that W110PD-L2 acts as an "elbow" that helps shorten PD-L2 engagement with PD-1 and therefore lower affinity. Furthermore, we identified a "latch" between the C and D β-strands of the binding face as the source of the PD-L2 affinity advantage. We show that the 3-fold affinity advantage of PD-L2 is the consequence of these two opposing features, the W110PD-L2 "elbow" and a C-D region "latch." Interestingly, using phylogenetic analysis, we found that these features evolved simultaneously upon the emergence of placental mammals, suggesting that PD-L2-affinity tuning was part of the alterations to the adaptive immune system required for placental gestation.
Keywords: PD-L2; T-cell biology; glycoprotein structure; immune checkpoint; immune receptors/ligands; immunotherapy; programmed cell death protein 1 (PD-1); programmed death ligand 1 (PD-L1); protein evolution; protein structure.
© 2020 Philips et al.
Conflict of interest statement
E. A. P., A. M., and X. P. K. are inventors on a pending patent application pertaining to this work
Figures
Comment in
-
A checkpoint cliffhanger at the dawn of placental mammals.J Biol Chem. 2020 Apr 3;295(14):4381-4382. doi: 10.1074/jbc.H120.013307. J Biol Chem. 2020. PMID: 32245902 Free PMC article.
References
-
- Freeman G. J., Long A. J., Iwai Y., Bourque K., Chernova T., Nishimura H., Fitz L. J., Malenkovich N., Okazaki T., Byrne M. C., Horton H. F., Fouser L., Carter L., Ling V., Bowman M. R., et al. (2000) Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J. Exp. Med. 192, 1027–1034 10.1084/jem.192.7.1027 - DOI - PMC - PubMed
-
- Latchman Y., Wood C. R., Chernova T., Chaudhary D., Borde M., Chernova I., Iwai Y., Long A. J., Brown J. A., Nunes R., Greenfield E. A., Bourque K., Boussiotis V. A., Carter L. L., Carreno B. M., et al. (2001) PD-L2 is a second ligand for PD-1 and inhibits T cell activation. Nat. Immunol. 2, 261–268 10.1038/85330 - DOI - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
