

Saltatory Conduction along Myelinated Axons Involves a Periaxonal Nanocircuit
- PMID: 31883793
- PMCID: PMC6978798
- DOI: 10.1016/j.cell.2019.11.039
Saltatory Conduction along Myelinated Axons Involves a Periaxonal Nanocircuit
Abstract
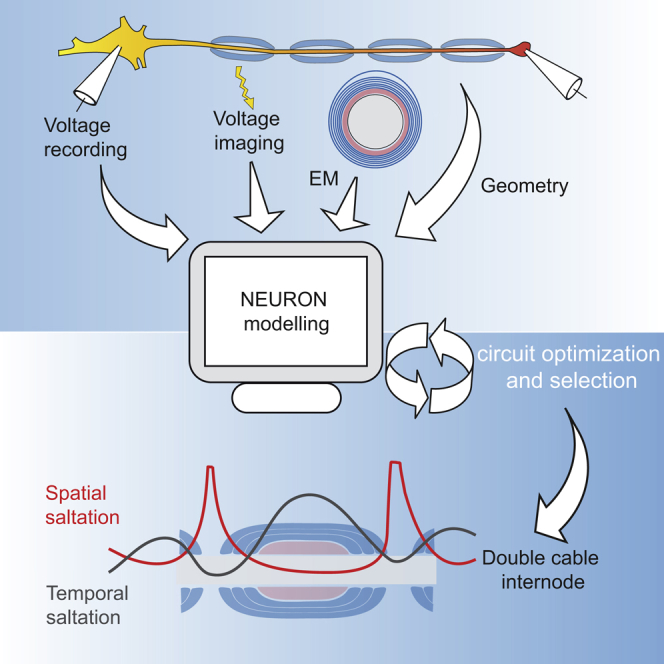
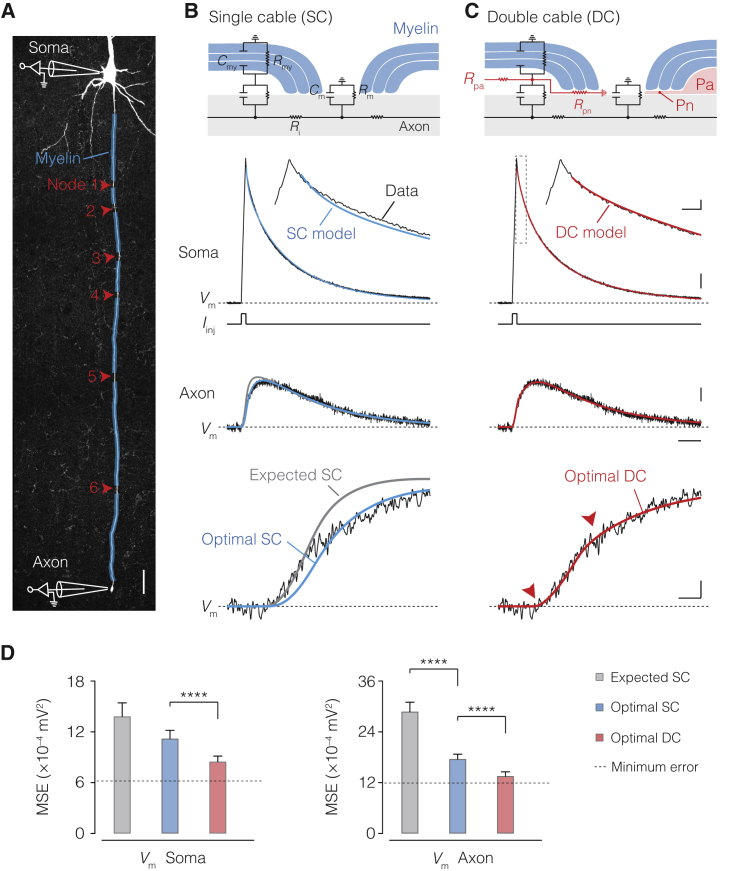
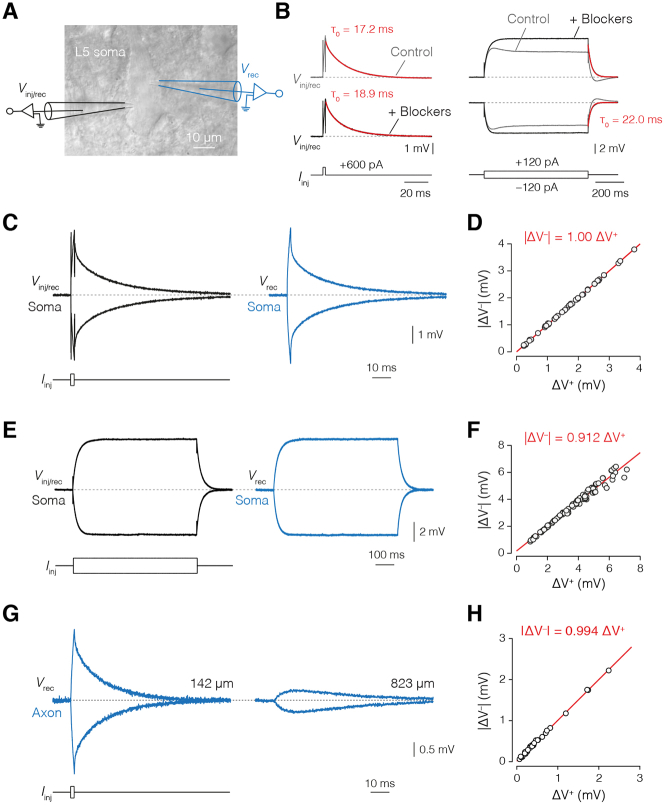
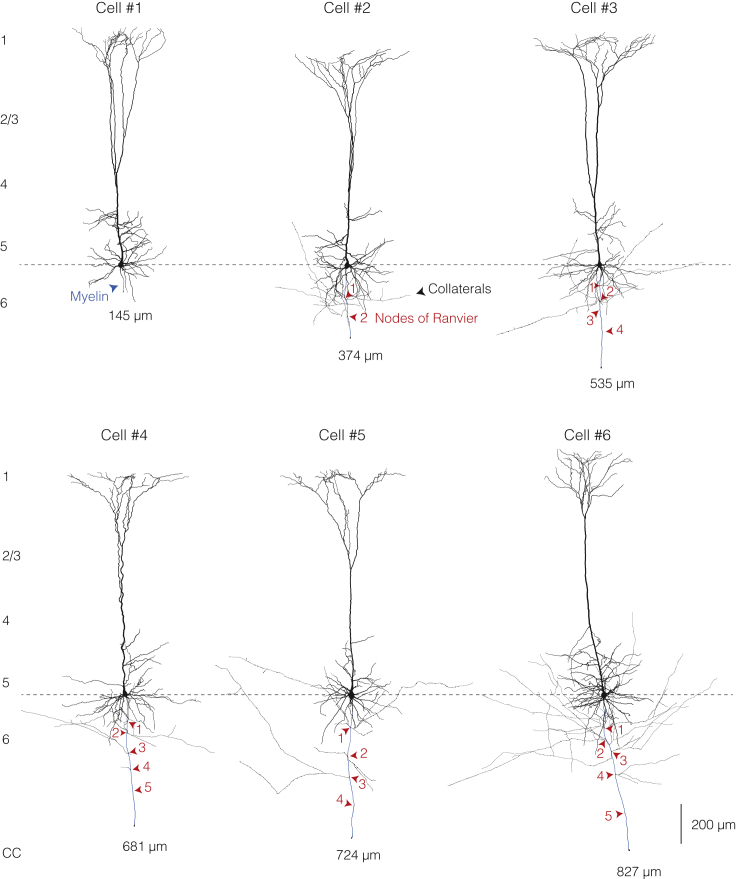
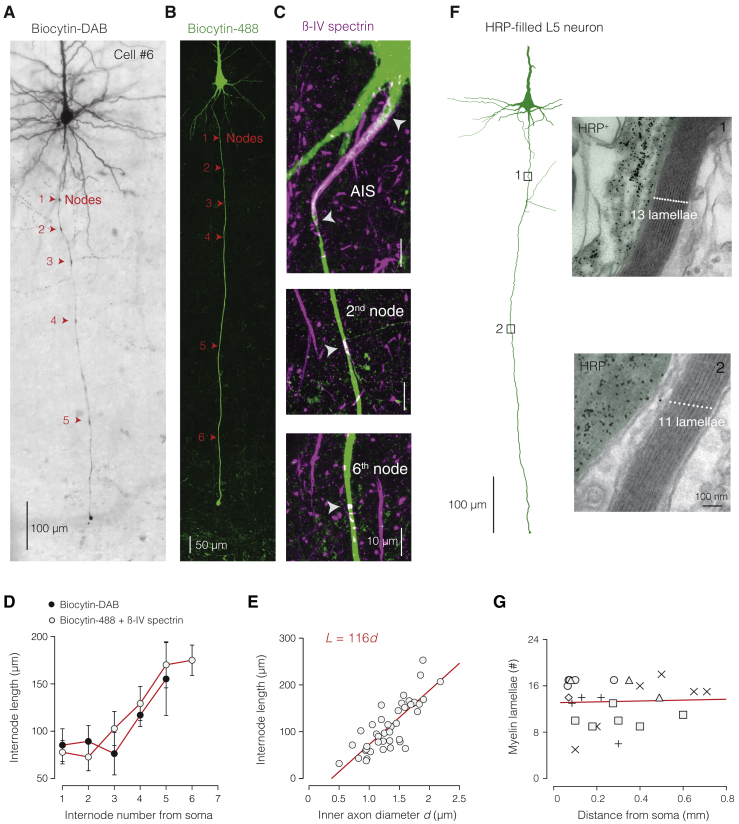
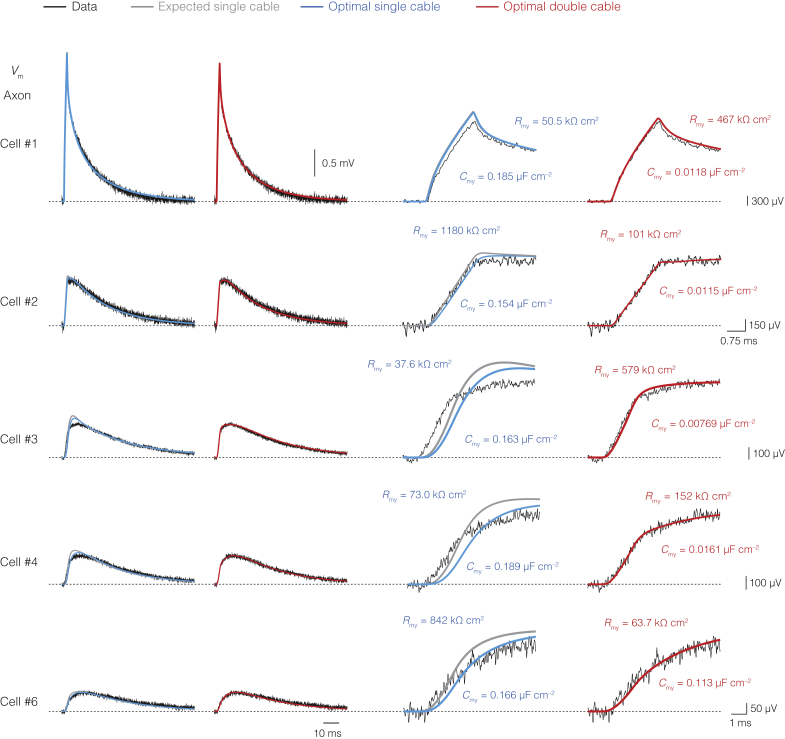
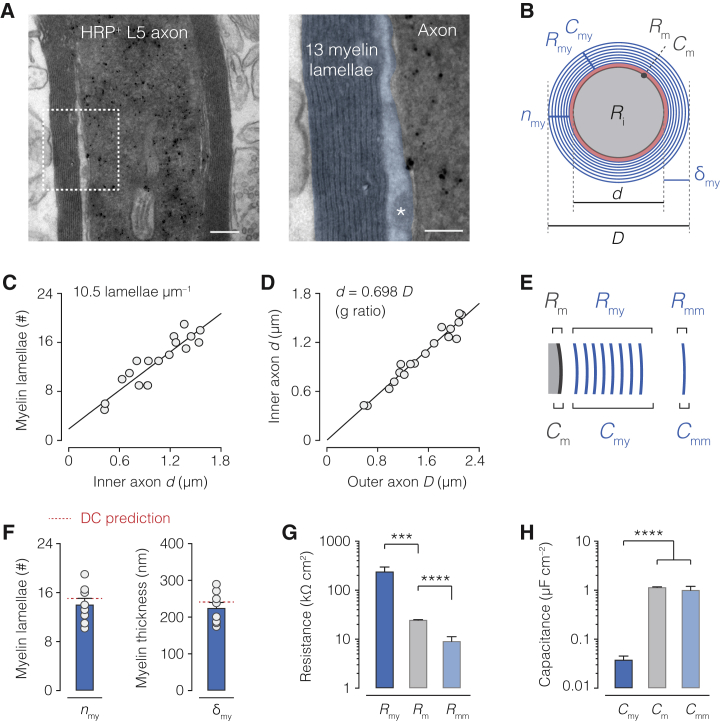
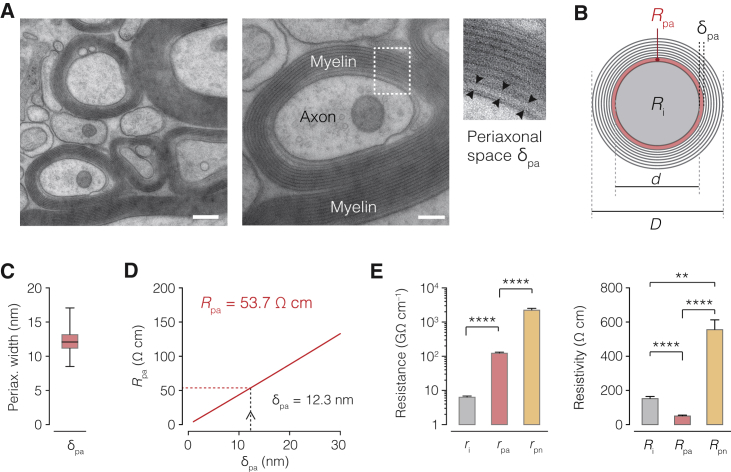
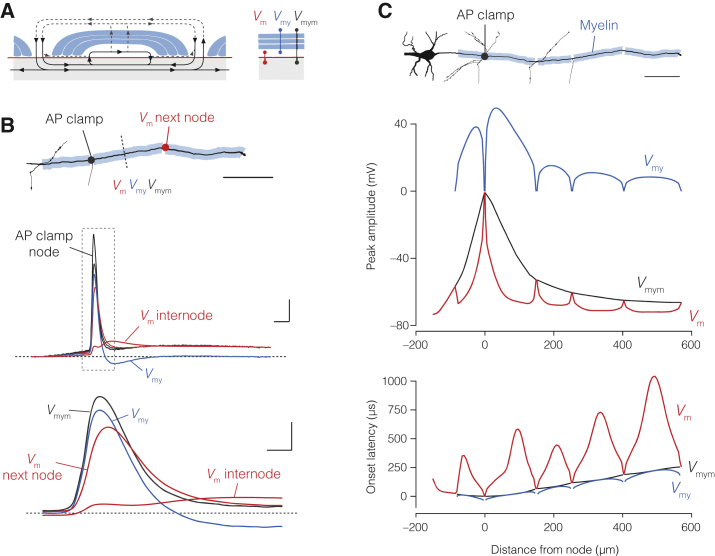
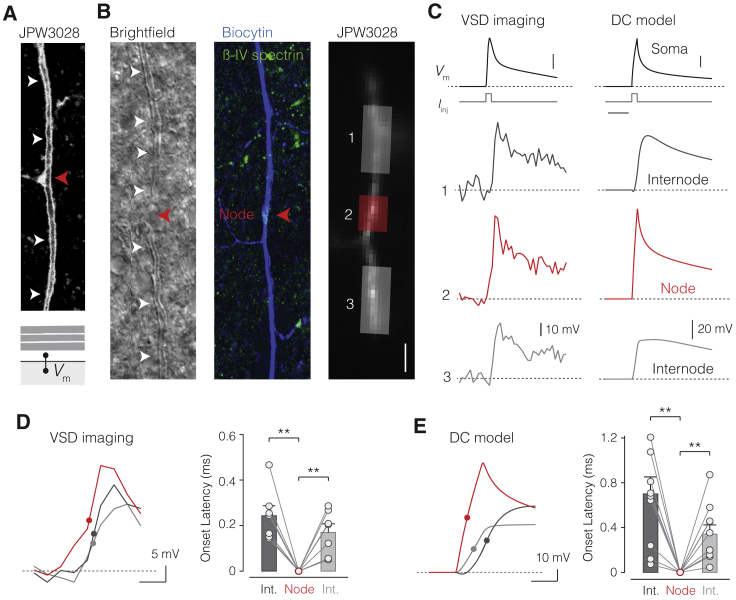
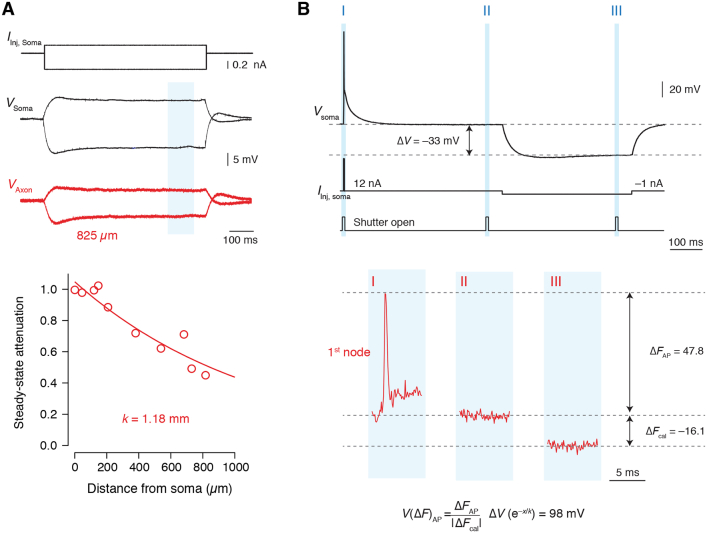
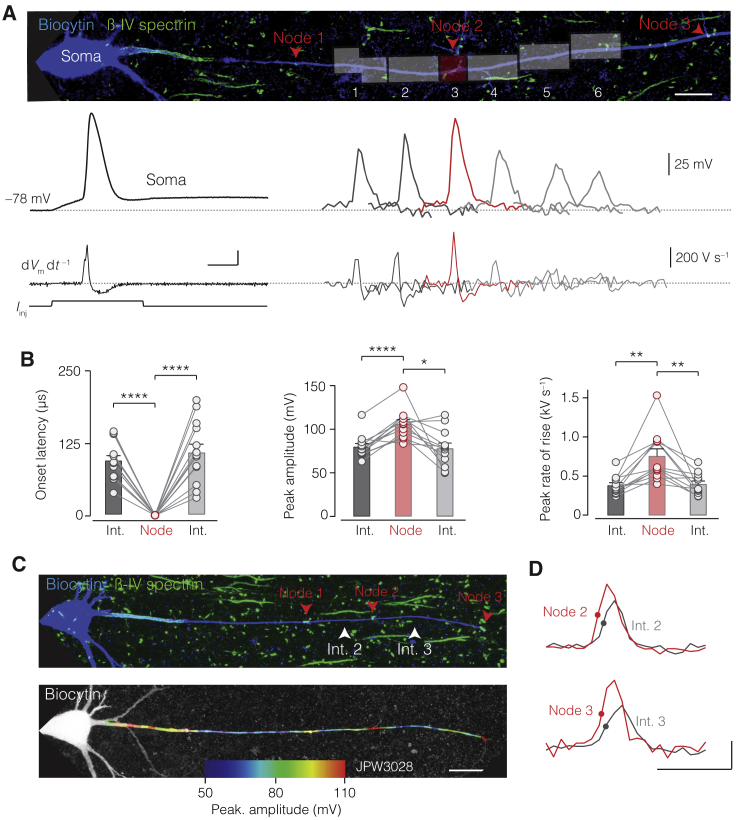
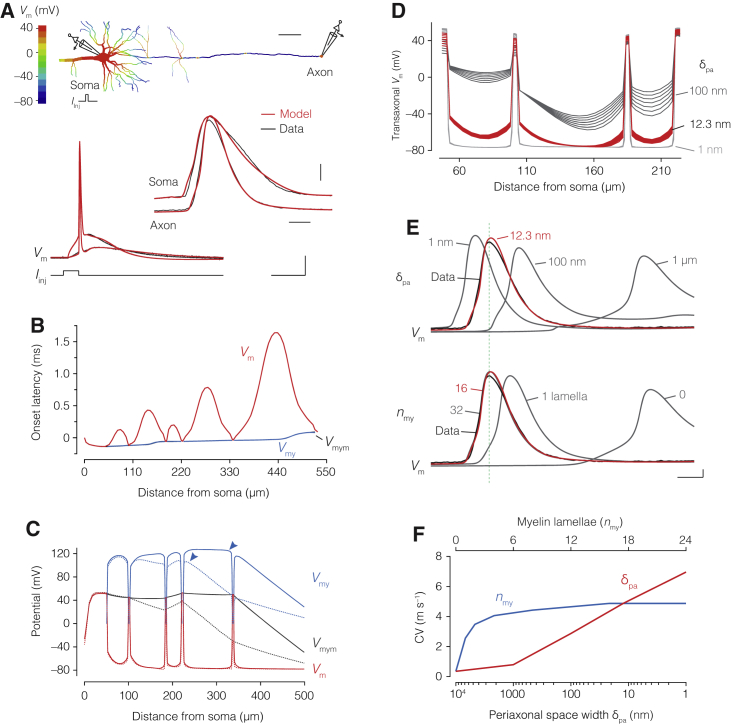
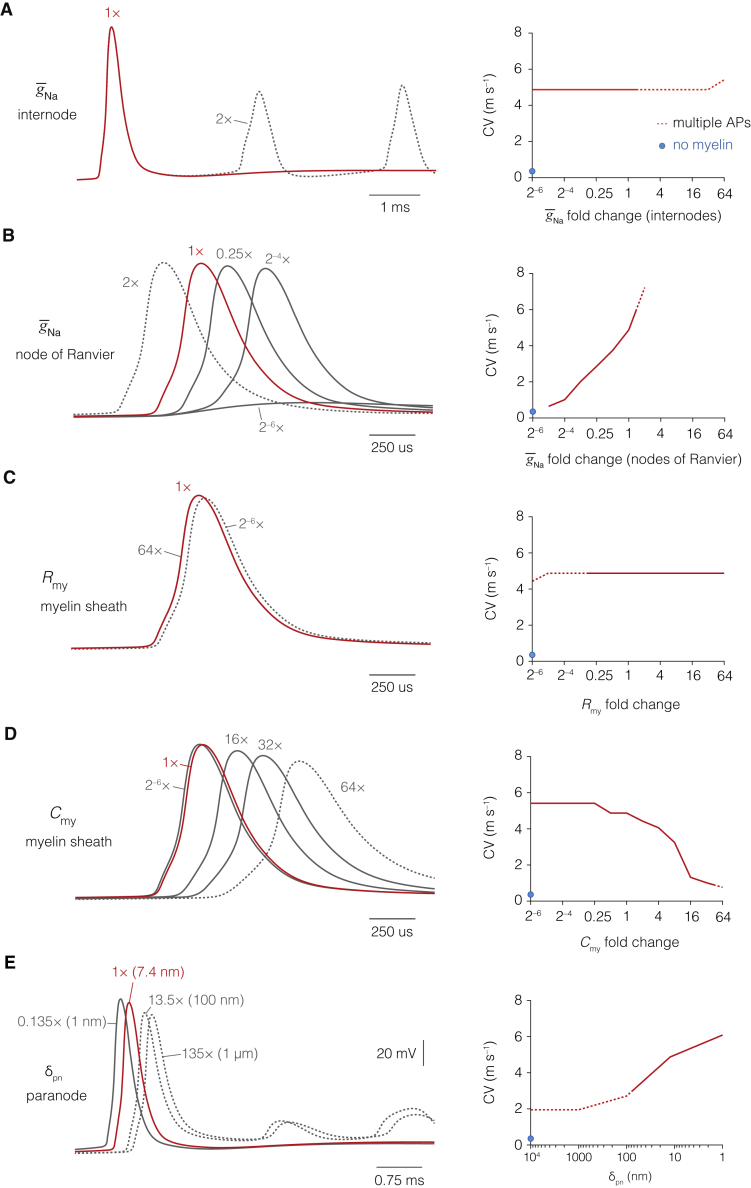
The propagation of electrical impulses along axons is highly accelerated by the myelin sheath and produces saltating or "jumping" action potentials across internodes, from one node of Ranvier to the next. The underlying electrical circuit, as well as the existence and role of submyelin conduction in saltatory conduction remain, however, elusive. Here, we made patch-clamp and high-speed voltage-calibrated optical recordings of potentials across the nodal and internodal axolemma of myelinated neocortical pyramidal axons combined with electron microscopy and experimentally constrained cable modeling. Our results reveal a nanoscale yet conductive periaxonal space, incompletely sealed at the paranodes, which separates the potentials across the low-capacitance myelin sheath and internodal axolemma. The emerging double-cable model reproduces the recorded evolution of voltage waveforms across nodes and internodes, including rapid nodal potentials traveling in advance of attenuated waves in the internodal axolemma, revealing a mechanism for saltation across time and space.
Keywords: action potential; axon; circuit; computational modelling; double cable; internode; myelin; periaxonal space; saltatory conduction; single cable.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Saltatory Conduction: Jumping to New Conclusions.Curr Biol. 2020 Apr 6;30(7):R326-R328. doi: 10.1016/j.cub.2020.02.037. Curr Biol. 2020. PMID: 32259510
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
