

Positioning Heterochromatin at the Nuclear Periphery Suppresses Histone Turnover to Promote Epigenetic Inheritance
- PMID: 31883795
- PMCID: PMC7102895
- DOI: 10.1016/j.cell.2019.12.004
Positioning Heterochromatin at the Nuclear Periphery Suppresses Histone Turnover to Promote Epigenetic Inheritance
Abstract
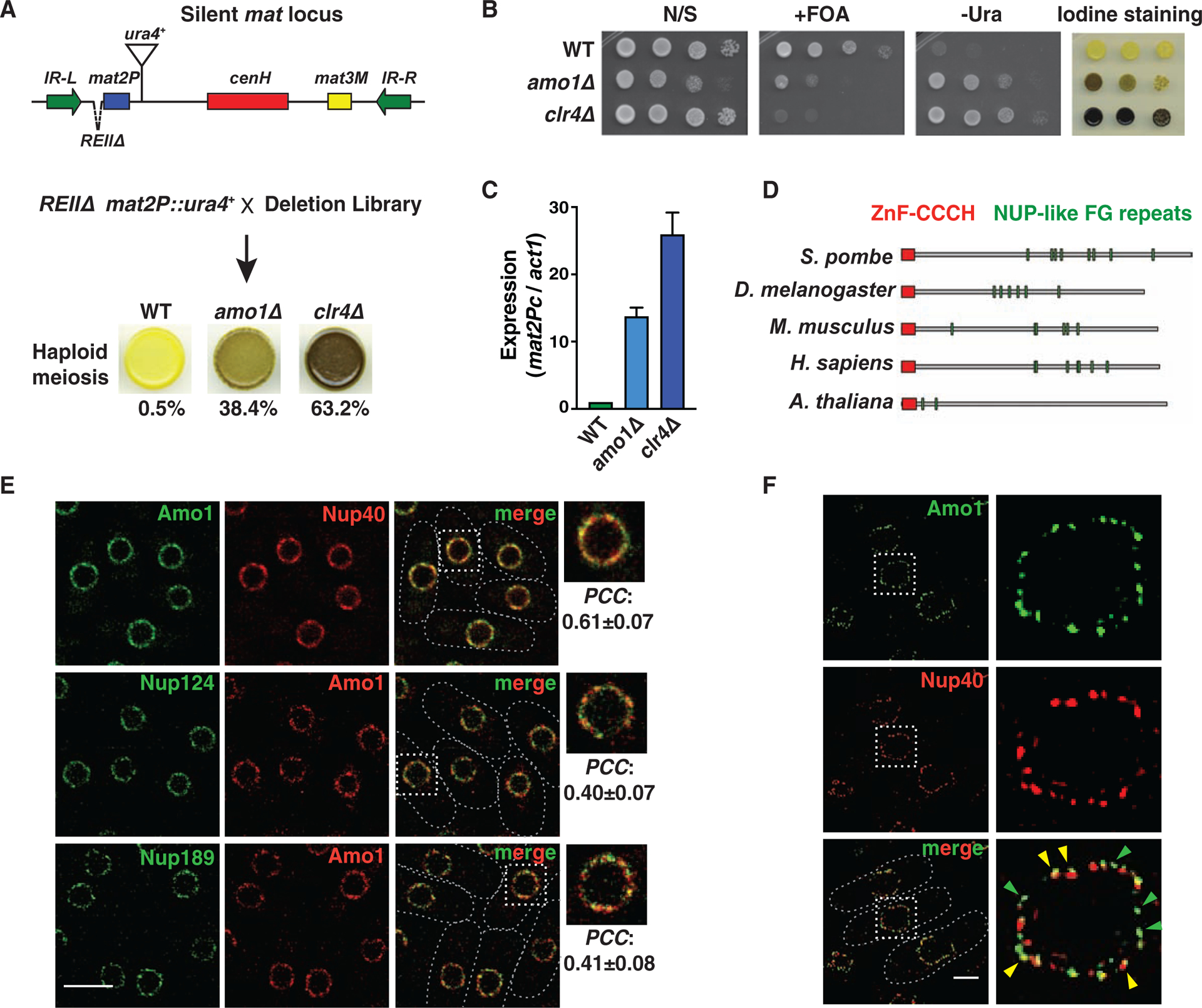
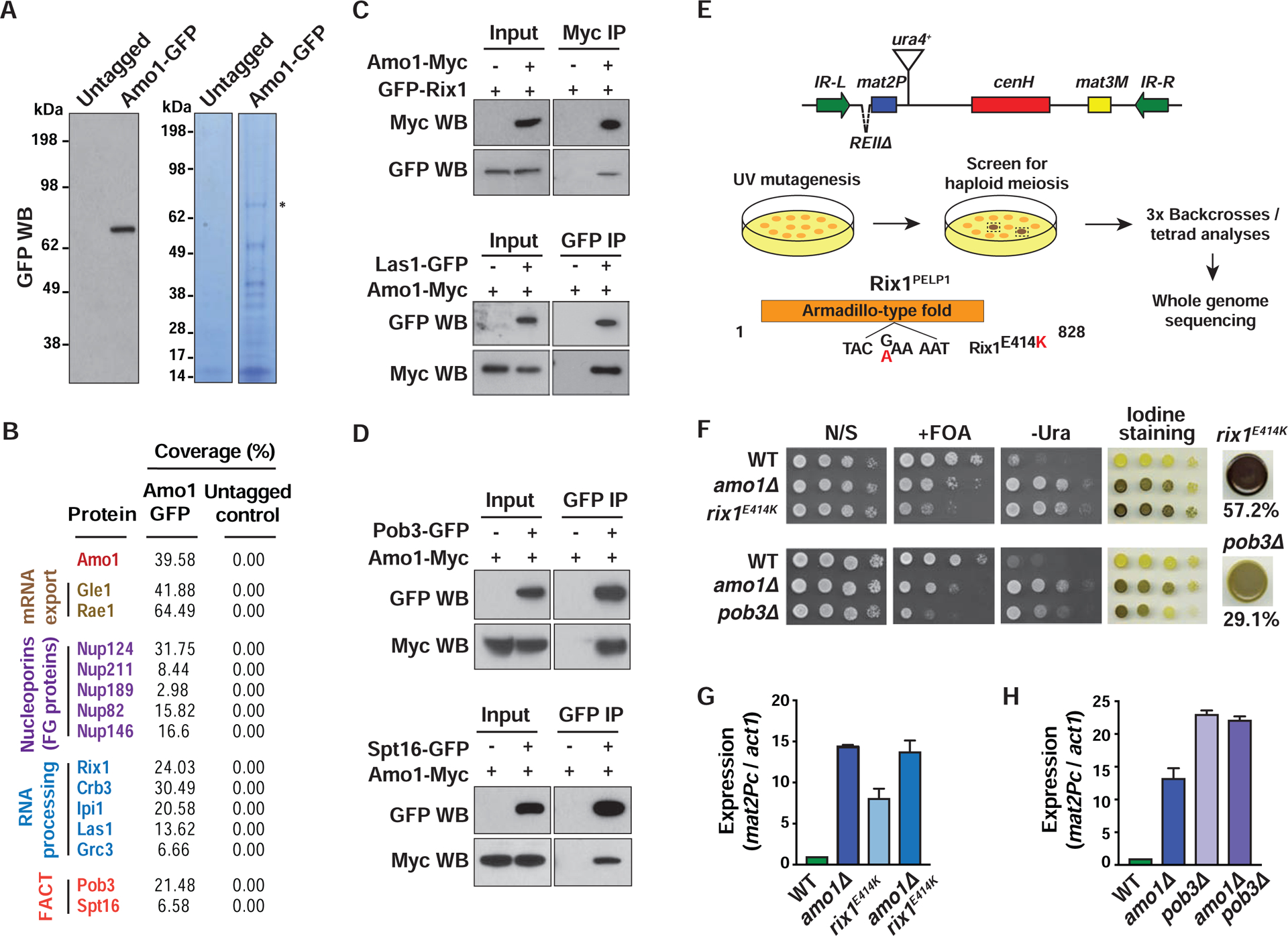
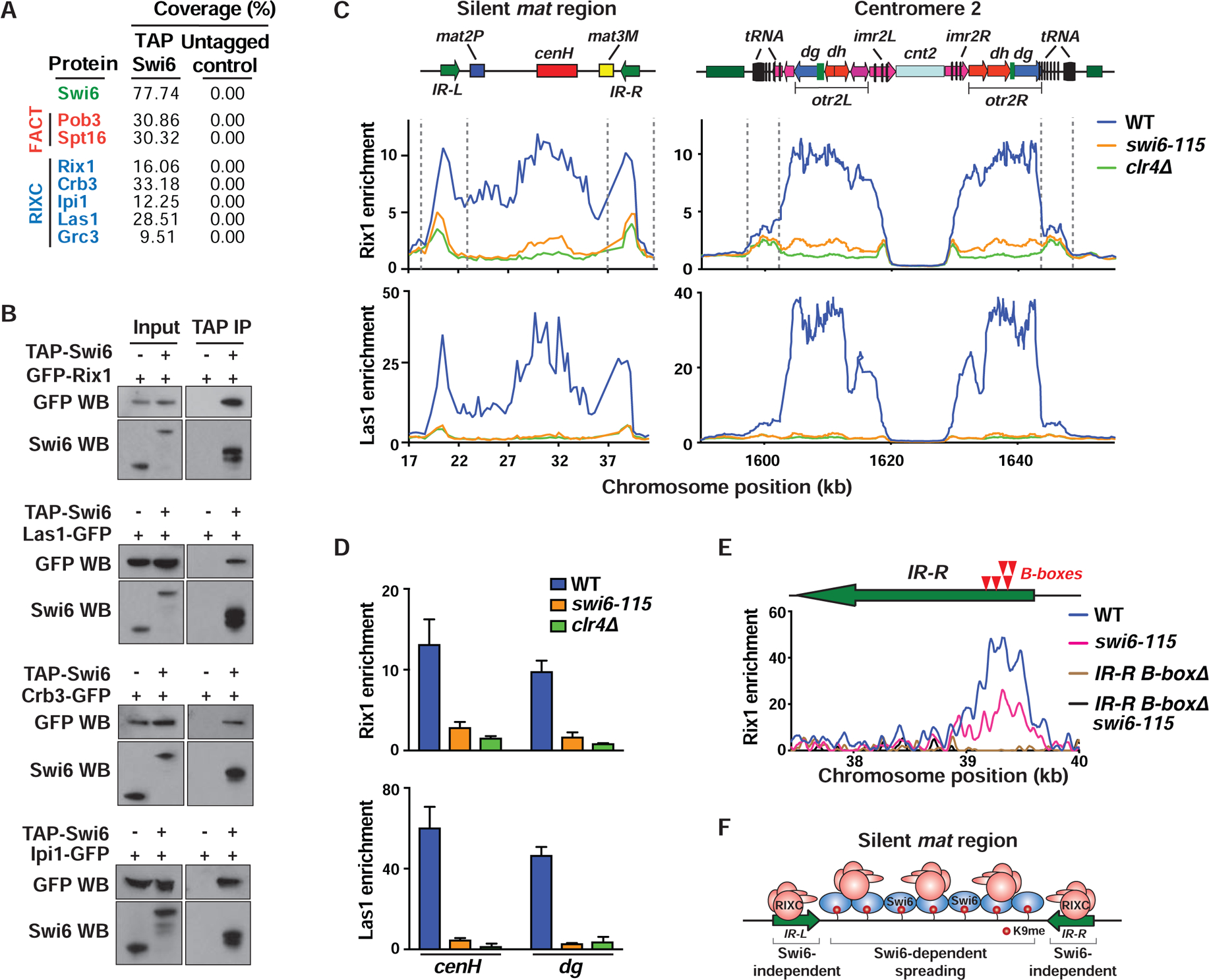
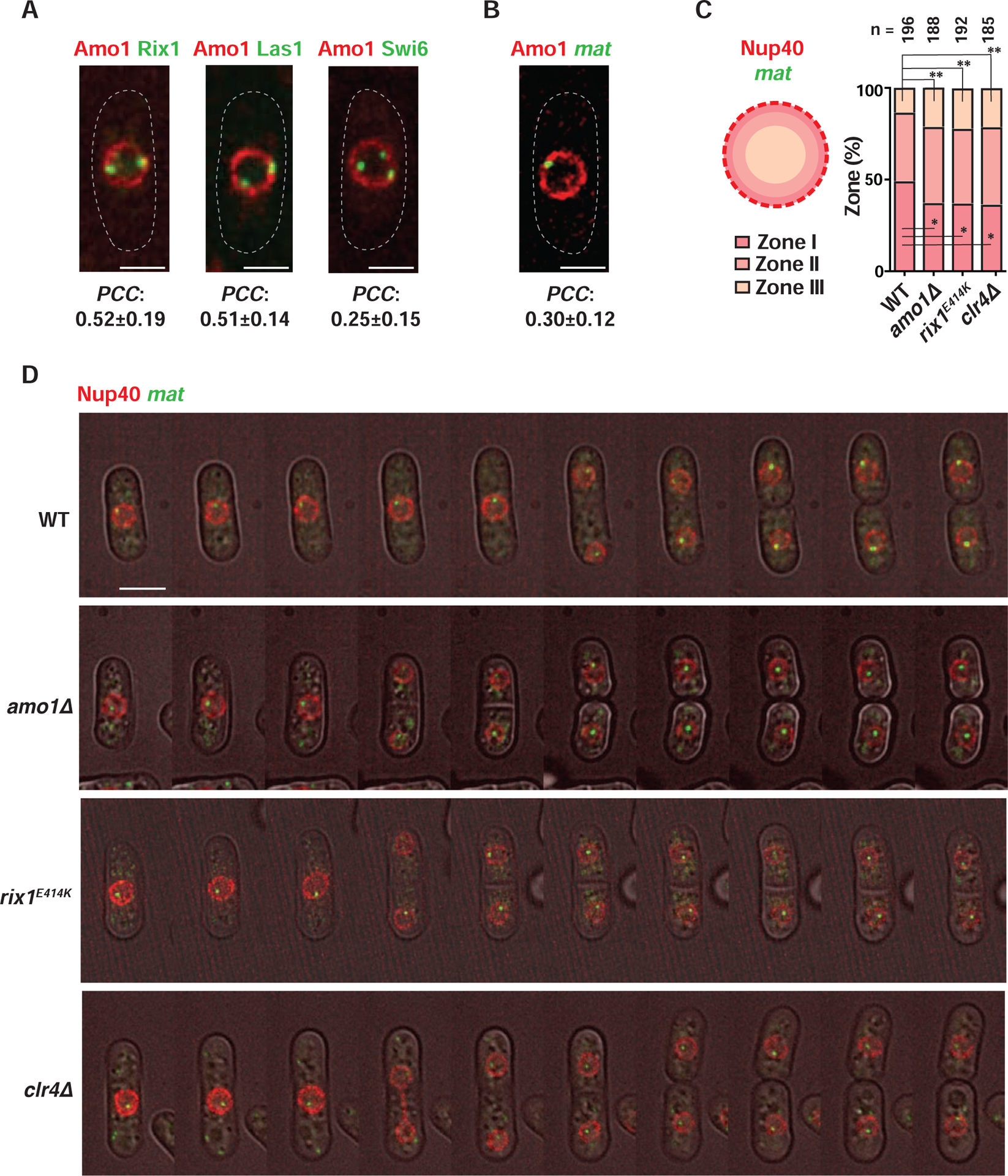
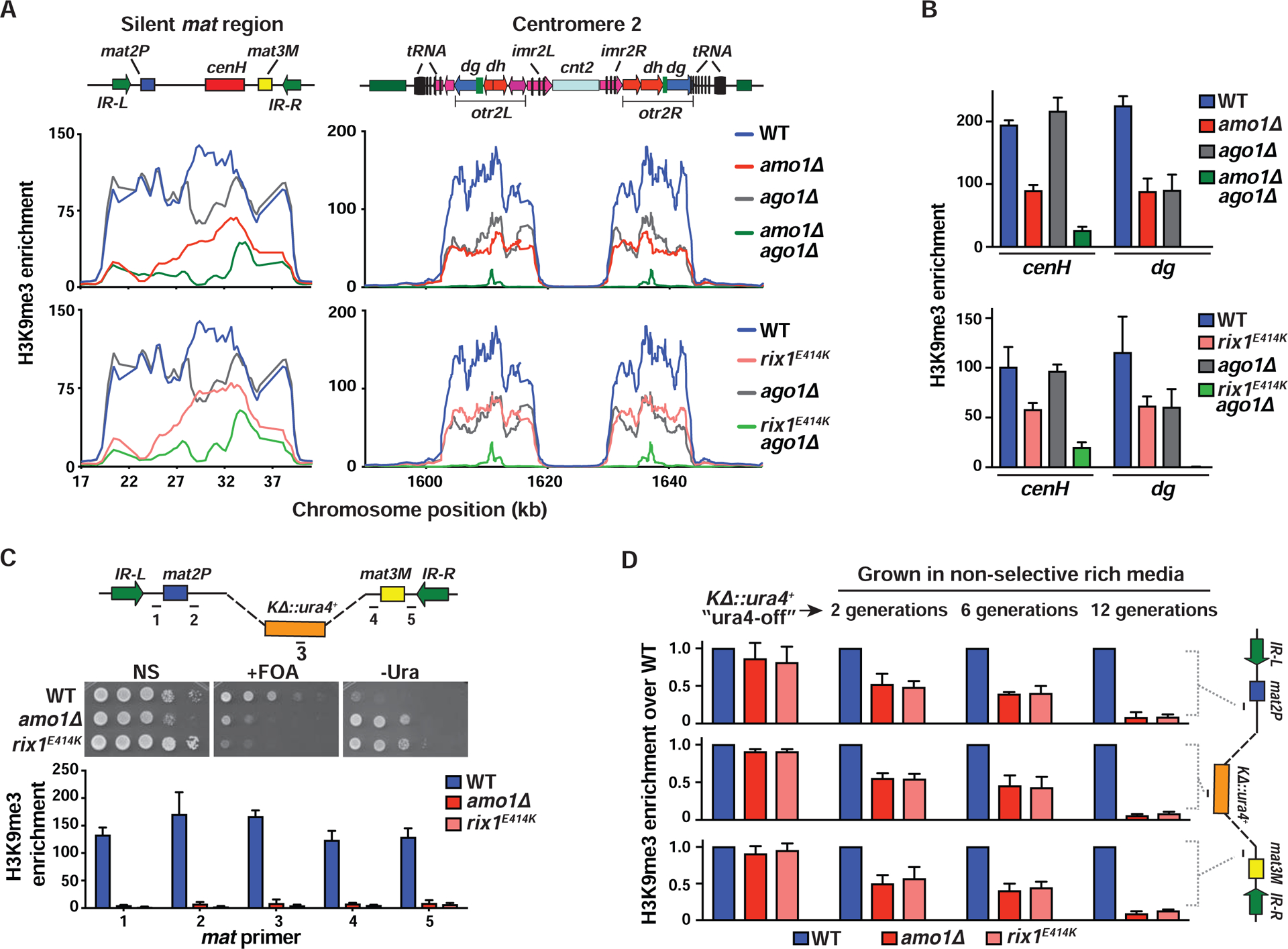
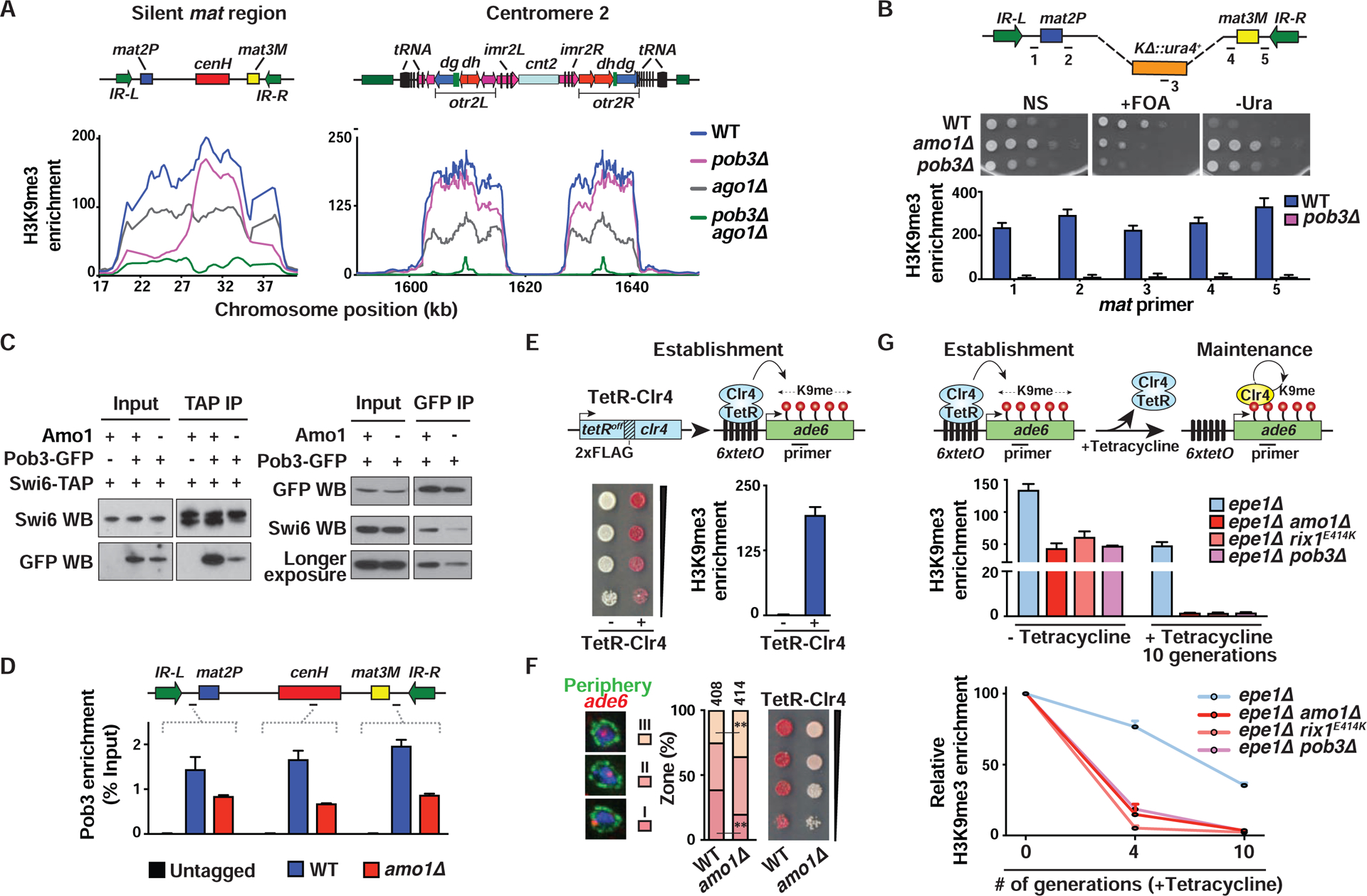
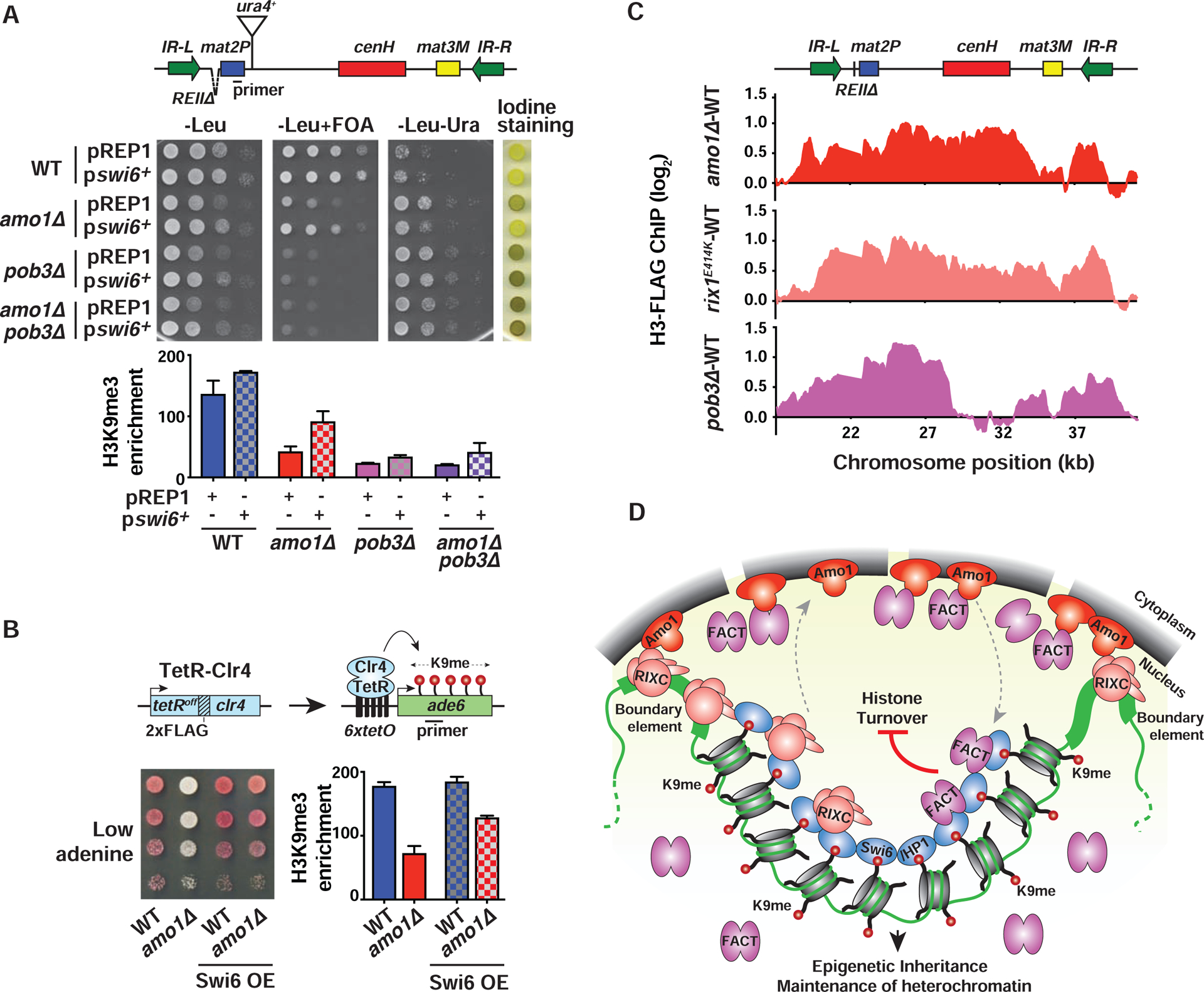
In eukaryotes, heterochromatin is generally located at the nuclear periphery. This study investigates the biological significance of perinuclear positioning for heterochromatin maintenance and gene silencing. We identify the nuclear rim protein Amo1NUPL2 as a factor required for the propagation of heterochromatin at endogenous and ectopic sites in the fission yeast genome. Amo1 associates with the Rix1PELP1-containing RNA processing complex RIXC and with the histone chaperone complex FACT. RIXC, which binds to heterochromatin protein Swi6HP1 across silenced chromosomal domains and to surrounding boundary elements, connects heterochromatin with Amo1 at the nuclear periphery. In turn, the Amo1-enriched subdomain is critical for Swi6 association with FACT that precludes histone turnover to promote gene silencing and preserve epigenetic stability of heterochromatin. In addition to uncovering conserved factors required for perinuclear positioning of heterochromatin, these analyses elucidate a mechanism by which a peripheral subdomain enforces stable gene repression and maintains heterochromatin in a heritable manner.
Keywords: chromatin; epigenetic inheritance; gene regulation; heterochromatin; heterochromatin spreading; histone methylation; nuclear organization; nuclear periphery.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
References
-
- Akhtar A, and Gasser SM (2007). The nuclear envelope and transcriptional control. Nat. Rev. Genet 8, 507–517. - PubMed
-
- Alfredsson-Timmins J, Henningson F, and Bjerling P (2007). The Clr4 methyltransferase determines the subnuclear localization of the mating-type region in fission yeast. J. Cell Sci 120, 1935–1943. - PubMed
-
- Andrulis ED, Neiman AM, Zappulla DC, and Sternglanz R (1998). Perinuclear localization of chromatin facilitates transcriptional silencing. Nature 394, 592–595. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
