

The steering gaits of sperm
- PMID: 31884910
- PMCID: PMC7017342
- DOI: 10.1098/rstb.2019.0149
The steering gaits of sperm
Abstract
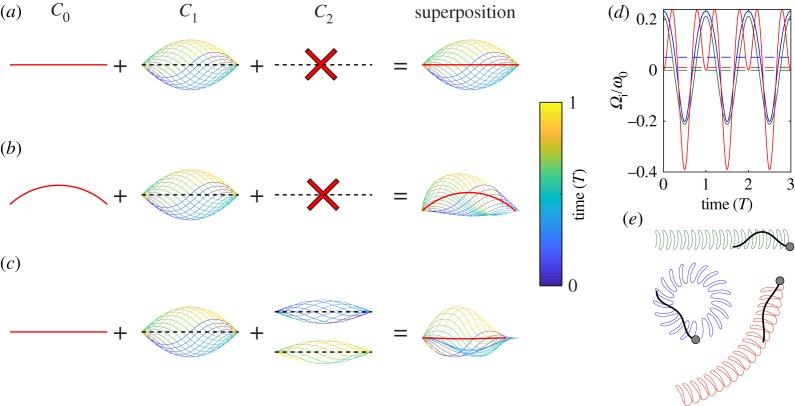
Sperm are highly specialized cells, which have been subject to substantial evolutionary pressure. Whereas some sperm features are highly conserved, others have undergone major modifications. Some of these variations are driven by adaptation to mating behaviours or fitness at the organismic level. Others represent alternative solutions to the same task. Sperm must find the egg for fertilization. During this task, sperm rely on long slender appendages termed flagella that serve as sensory antennas, propellers and steering rudders. The beat of the flagellum is periodic. The resulting travelling wave generates the necessary thrust for propulsion in the fluid. Recent studies reveal that, for steering, different species rely on different fundamental features of the beat wave. Here, we discuss some examples of unity and diversity across sperm from different species with a particular emphasis on the steering mechanisms. This article is part of the Theo Murphy meeting issue 'Unity and diversity of cilia in locomotion and transport'.
Keywords: chirality; cilia; cytoskeleton; navigation; symmetry-breaking.
Conflict of interest statement
We declare we have no competing interests.
Figures



References
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
