

Cyanidioschyzon merolae aurora kinase phosphorylates evolutionarily conserved sites on its target to regulate mitochondrial division
- PMID: 31886415
- PMCID: PMC6925296
- DOI: 10.1038/s42003-019-0714-x
Cyanidioschyzon merolae aurora kinase phosphorylates evolutionarily conserved sites on its target to regulate mitochondrial division
Abstract
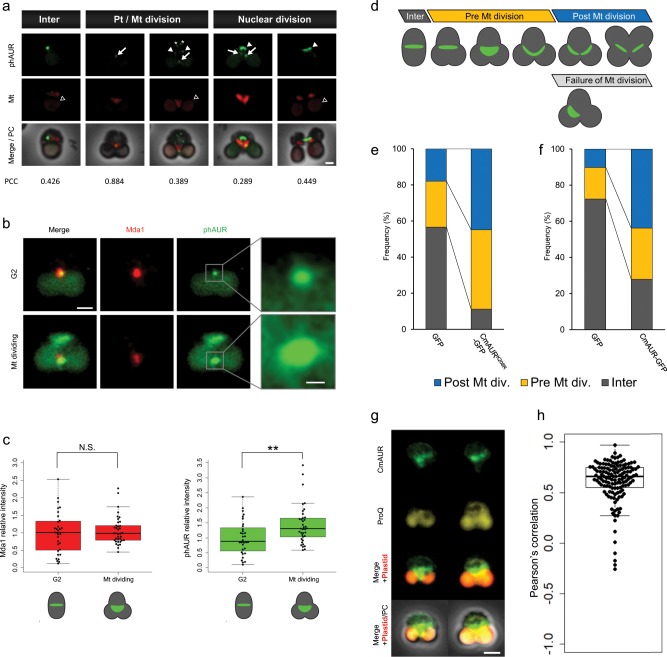
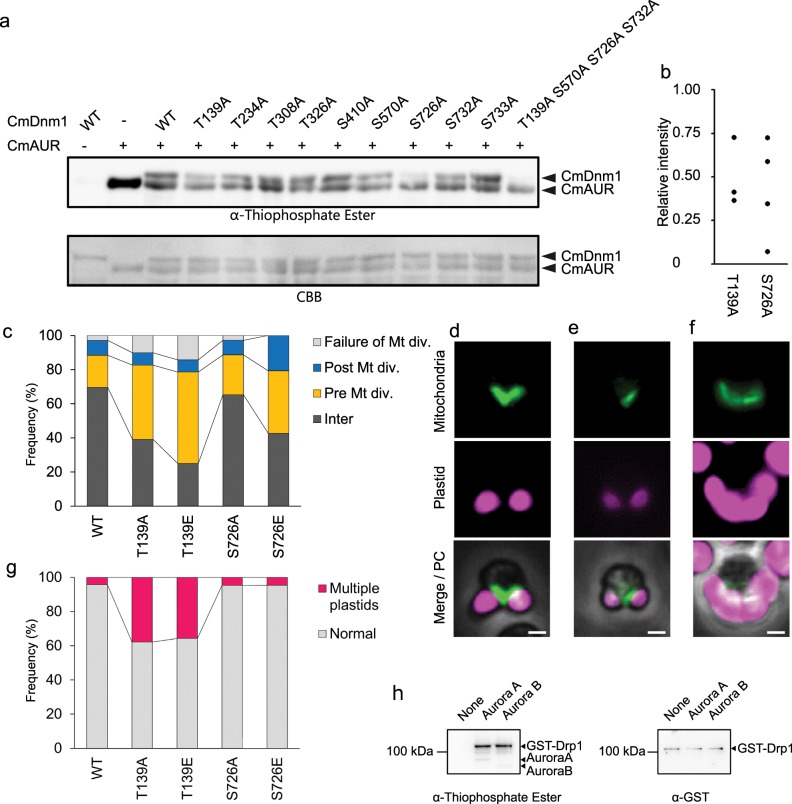
The mitochondrion is an organelle that was derived from an endosymbiosis. Although regulation of mitochondrial growth by the host cell is necessary for the maintenance of mitochondria, it is unclear how this regulatory mechanism was acquired. To address this, we studied the primitive unicellular red alga Cyanidioschyzon merolae, which has the simplest eukaryotic genome and a single mitochondrion. Here we show that the C. merolae Aurora kinase ortholog CmAUR regulates mitochondrial division through phosphorylation of mitochondrial division ring components. One of the components, the Drp1 ortholog CmDnm1, has at least four sites phosphorylated by CmAUR. Depletion of the phosphorylation site conserved among eukaryotes induced defects such as mitochondrial distribution on one side of the cell. Taken together with the observation that human Aurora kinase phosphorylates Drp1 in vitro, we suggest that the phosphoregulation is conserved from the simplest eukaryotes to mammals, and was acquired at the primitive stage of endosymbiosis.
Keywords: Cell biology; Plant sciences.
© The Author(s) 2019.
Conflict of interest statement
Competing interestsThe authors declare no competing interests.
Figures


Similar articles
-
The kinesin-like protein TOP promotes Aurora localisation and induces mitochondrial, chloroplast and nuclear division.J Cell Sci. 2013 Jun 1;126(Pt 11):2392-400. doi: 10.1242/jcs.116798. Epub 2013 Apr 2. J Cell Sci. 2013. PMID: 23549784
-
Dynamic recruitment of dynamin for final mitochondrial severance in a primitive red alga.Proc Natl Acad Sci U S A. 2003 Feb 18;100(4):2146-51. doi: 10.1073/pnas.0436886100. Epub 2003 Feb 3. Proc Natl Acad Sci U S A. 2003. PMID: 12566569 Free PMC article.
-
Simple prerequisite of presequence for mitochondrial protein import in the unicellular red alga Cyanidioschyzon merolae.J Cell Sci. 2024 Jul 15;137(14):jcs262042. doi: 10.1242/jcs.262042. Epub 2024 Jul 23. J Cell Sci. 2024. PMID: 38940185 Free PMC article.
-
Structure, function and evolution of the mitochondrial division apparatus.Biochim Biophys Acta. 2006 May-Jun;1763(5-6):510-21. doi: 10.1016/j.bbamcr.2006.03.007. Epub 2006 Apr 5. Biochim Biophys Acta. 2006. PMID: 16690143 Review.
-
Aurora Kinases Throughout Plant Development.Trends Plant Sci. 2016 Jan;21(1):69-79. doi: 10.1016/j.tplants.2015.10.001. Epub 2015 Nov 23. Trends Plant Sci. 2016. PMID: 26616196 Review.
Cited by
-
The hypervariable region of atlastin-1 is a site for intrinsic and extrinsic regulation.J Cell Biol. 2021 Nov 1;220(11):e202104128. doi: 10.1083/jcb.202104128. Epub 2021 Sep 21. J Cell Biol. 2021. PMID: 34546351 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
