

hnRNPA2B1 inhibits the exosomal export of miR-503 in endothelial cells
- PMID: 31894362
- PMCID: PMC11104873
- DOI: 10.1007/s00018-019-03425-6
hnRNPA2B1 inhibits the exosomal export of miR-503 in endothelial cells
Abstract
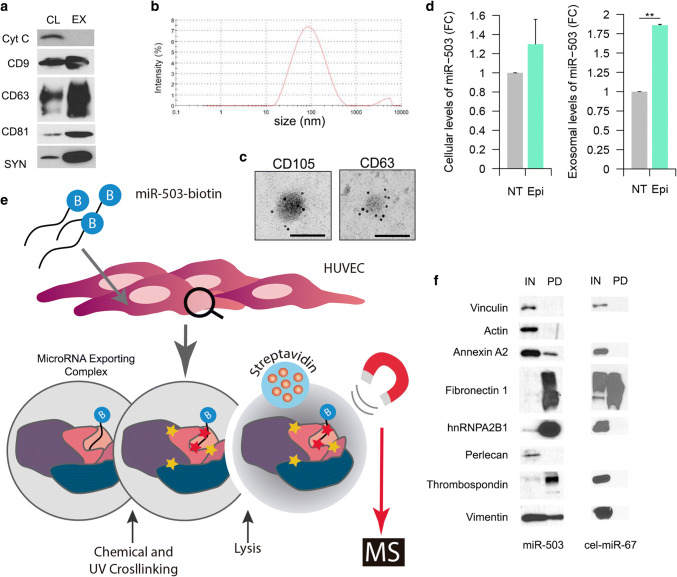
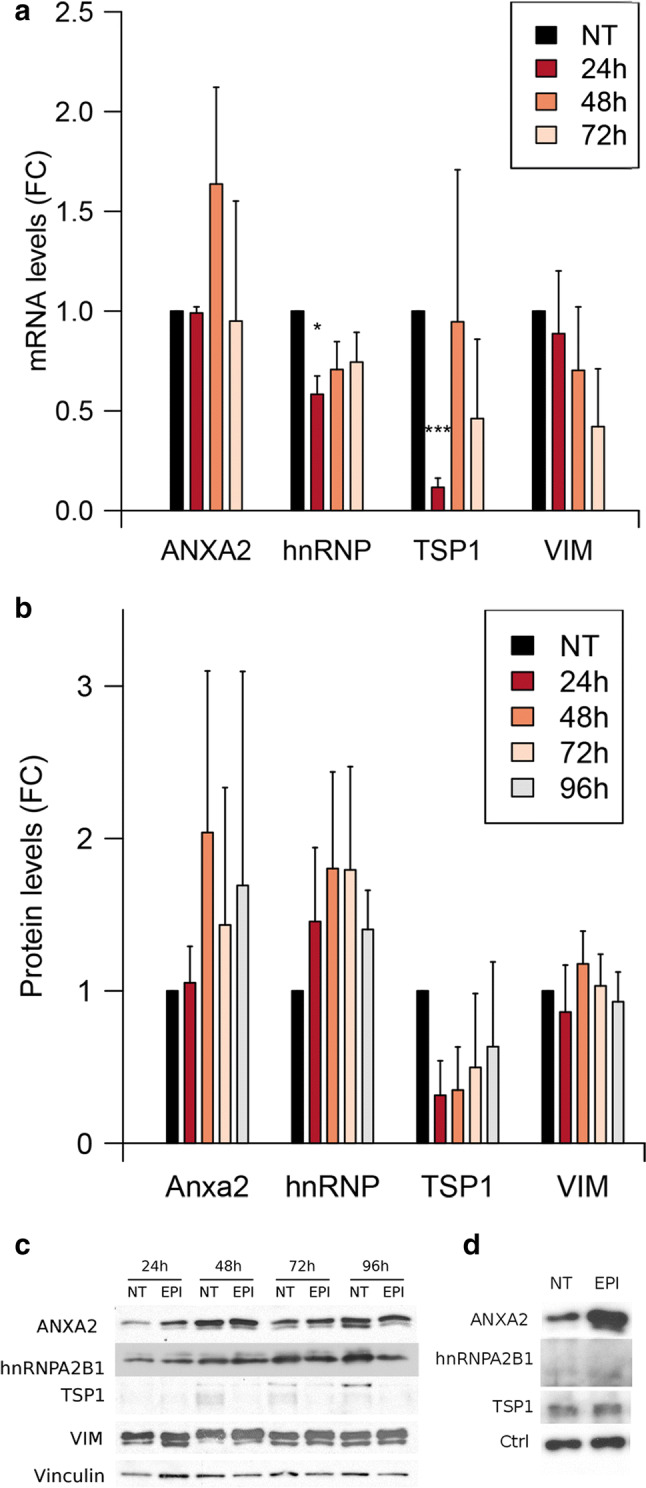
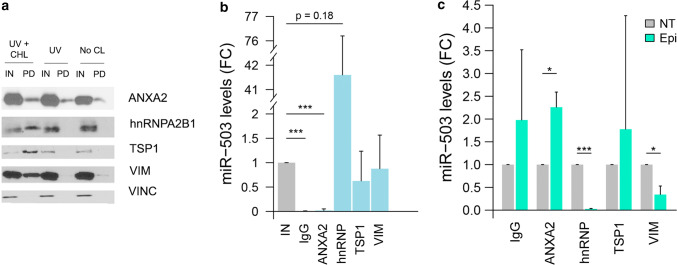
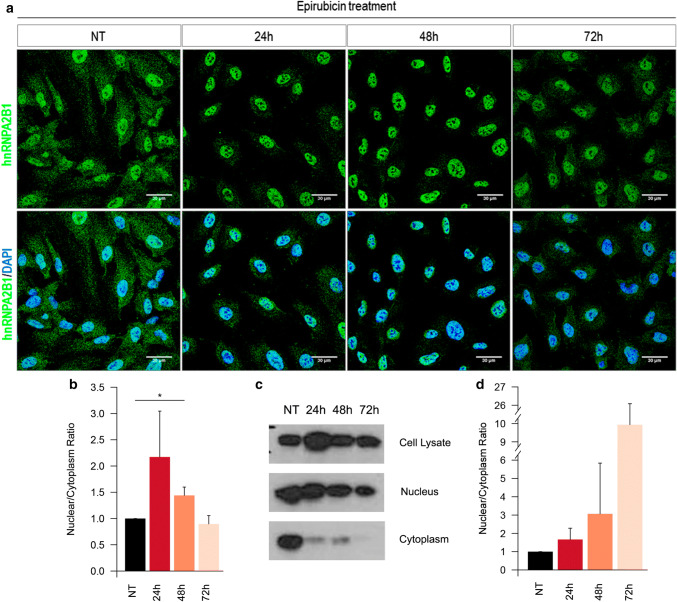
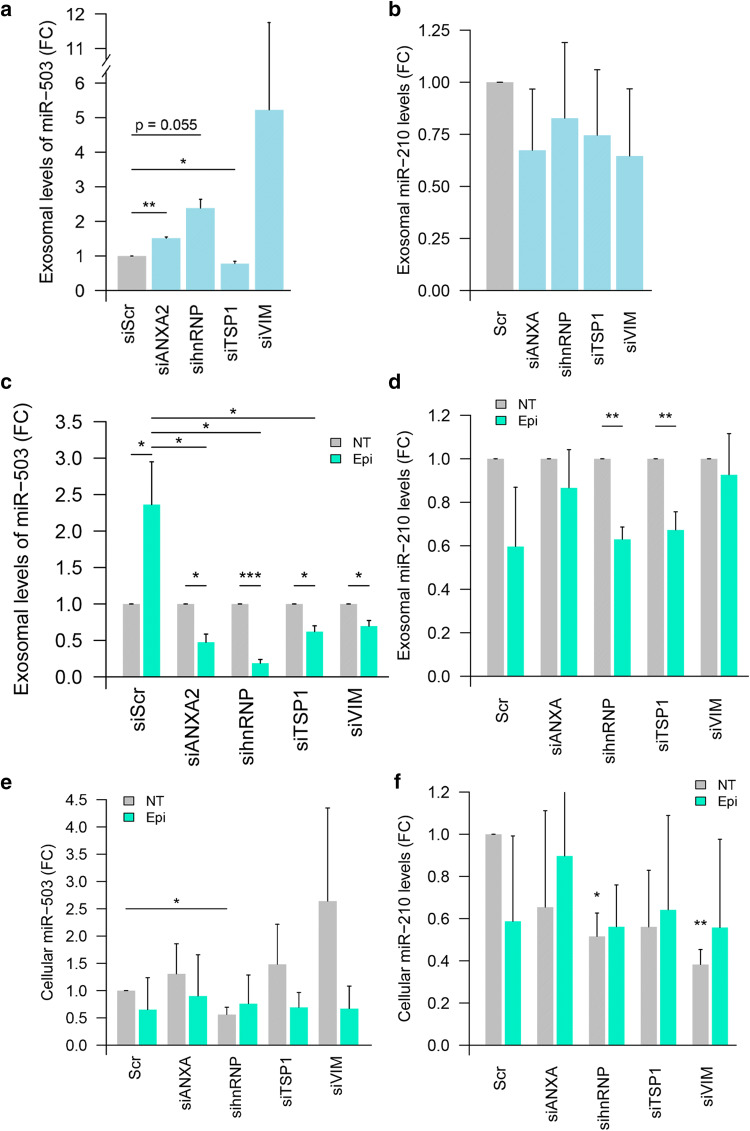
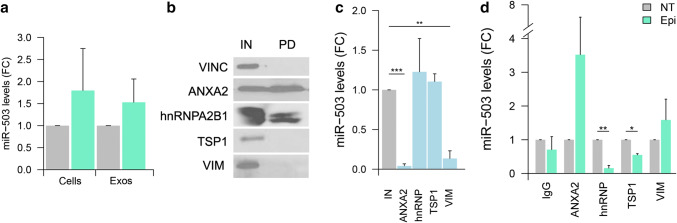
The chemotherapeutic drug epirubicin increases the exosomal export of miR-503 in endothelial cells. To understand the mechanisms behind this process, we transfected endothelial cells with miR-503 carrying a biotin tag. Then, we pulled-down the proteins interacting with miR-503 and studied their role in microRNA exosomal export. A total of four different binding partners were identified by mass spectrometry and validated by western blotting and negative controls, among them ANXA2 and hnRNPA2B1. Using knock-down systems combined with pull-down analysis, we determined that epirubicin mediates the export of miR-503 by disrupting the interaction between hnRNPA2B1 and miR-503. Then, both ANXA2 and miR-503 are sorted into exosomes while hnRNPA2B1 is relocated into the nucleus. The combination of these processes culminates in the increased export of miR-503. These results suggest, for the first time, that RNA-binding proteins can negatively regulate the exosomal sorting of microRNAs.
Keywords: EVs; Exosomal export; Exosomes; MicroRNAs; RNA-binding proteins.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
