

DNA double strand break repair in Escherichia coli perturbs cell division and chromosome dynamics
- PMID: 31895943
- PMCID: PMC6959608
- DOI: 10.1371/journal.pgen.1008473
DNA double strand break repair in Escherichia coli perturbs cell division and chromosome dynamics
Abstract
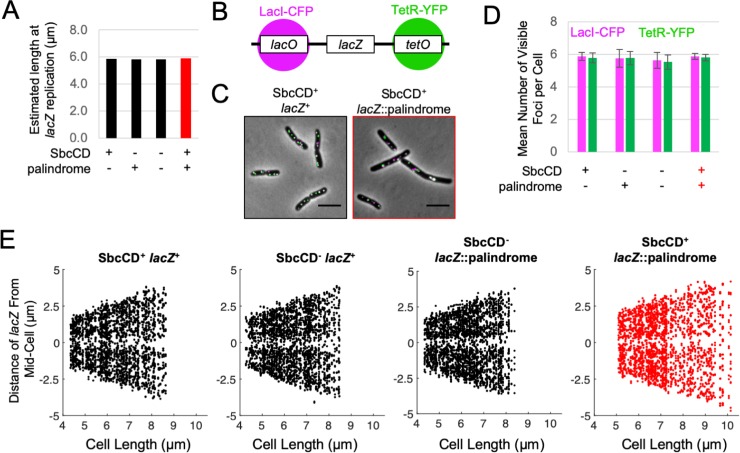
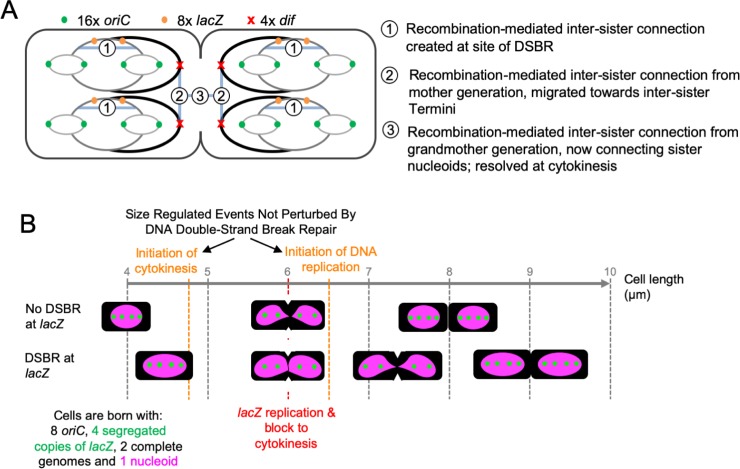
To prevent the transmission of damaged genomic material between generations, cells require a system for accommodating DNA repair within their cell cycles. We have previously shown that Escherichia coli cells subject to a single, repairable site-specific DNA double-strand break (DSB) per DNA replication cycle reach a new average cell length, with a negligible effect on population growth rate. We show here that this new cell size distribution is caused by a DSB repair-dependent delay in completion of cell division. This delay occurs despite unperturbed cell size regulated initiation of both chromosomal DNA replication and cell division. Furthermore, despite DSB repair altering the profile of DNA replication across the genome, the time required to complete chromosomal duplication is invariant. The delay in completion of cell division is accompanied by a DSB repair-dependent delay in individualization of sister nucleoids. We suggest that DSB repair events create inter-sister connections that persist until those chromosomes are separated by a closing septum.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
